Биосинтез холестерина его регуляция
Содержание статьи
Синтез холестерола должен быть согласован с его выведением
Синтез холестерола в организме составляет примерно 0,5-0,8 г/сут, примерно 50% образуется в печени, около 15% в кишечнике. Все клетки организма способны синтезировать холестерол. Поступление с пищей составляет около 0,4 г/сут.
Единственным реальным способом выведения холестерола является желчь – до 1 г/сут.
Биосинтез холестерола
Биосинтез холестерола происходит в эндоплазматическом ретикулуме. Источником всех атомов углерода в молекуле является ацетил-SКоА, поступающий сюда из митохондрий в составе цитрата, также как при синтезе жирных кислот. При биосинтезе холестерола затрачивается 18 молекул АТФ и 13 молекул НАДФН.
Образование холестерола идет более чем в 30 реакциях, которые можно сгруппировать в несколько этапов.
1. Синтез мевалоновой кислоты.
Первые две реакции синтеза совпадают с реакциями кетогенеза, но после синтеза 3-гидрокси-3-метилглутарил-SКоА вступает в действие фермент гидроксиметил-глутарил-SКоА-редуктаза (ГМГ-SКоА-редуктаза), образующая мевалоновую кислоту.
2. Синтез изопентенилдифосфата. На этом этапе при использовании АТФ мевалоновая кислота трижды фосфорилируется. Затем промежуточный продукт декарбоксилируется и дефосфорилируется с получением изопентенилдифосфата.
3. После объединения трех молекул изопентенилдифосфата (если не считать промежуточных превращений) синтезируется фарнезилдифосфат.
4. Синтез сквалена происходит при связывания двух остатков фарнезилдифосфата.
5. После сложных реакций линейный сквален циклизуется в ланостерол.
6. Удаление лишних метильных групп, восстановление и изомеризация молекулы приводит к появлению холестерола.
Регуляция синтеза
Регуляторным ферментом является гидроксиметилглутарил-SКоА-редуктаза, активность которой может изменяться в 100 и более раз.
1. Метаболическая регуляция – по принципу обратной отрицательной связи фермент аллостерически ингибируется конечным продуктом реакции – холестеролом. Это помогает поддерживать внутриклеточное содержание холестерола постоянным.
2. Регуляция транскрипции гена ГМГ-SКоА-редуктазы (в печени) – холестерол и желчные кислоты подавляют считывание гена и уменьшают количество фермента.
3. Ковалентная модификация при гормональной регуляции:
- Инсулин, активируя протеин-фосфатазу, способствует переходу фермента в активное состояние.
Важным следствием такой регуляции является активация кетогенеза при отсутствии инсулина.
- Глюкагон и адреналин посредством аденилатциклазного механизма активируют протеинкиназу А, которая фосфорилирует фермент и переводит его в неактивную форму.
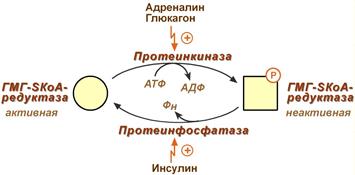
Регуляция активности гидроксиметилглутарил-S-КоА-редуктазы
Кроме указанных гормонов, на ГМГ-SКоА-редуктазу действуют эстрогены и тиреоидные гормоны (повышают активность) и глюкокортикоиды (снижают активность).
Изменение транскрипции гена ГМГ-КоА редуктазы (генетическая регуляция) осуществляется стерол-регулируемым элементом в ДНК (SREBP, sterol regulatory element-binding protein) с которым способны связываться белки — факторы SREBP. Эти факторы при достаточном количестве холестерина в клетке закреплены в мембране ЭПР. Когда уровень холестерина падает, факторы SREBP активируются под действием специфичных протеаз комплекса Гольджи, передвигаются в ядро, взаимодействуют на ДНК с участком SREBP и стимулируют биосинтез холестерина.
Скорость биосинтеза холестерола также зависит от концентрации специфического белка-переносчика, обеспечивающего связывание и транспорт гидрофобных промежуточных метаболитов синтеза.
Источник
Вопрос 54. Биосинтез холестерина и его этапы. Регуляция.
Реакции
синтеза холестерола происходят в
цитозоле клеток. Это один из самых
длинных метаболических путей в организме
человека.
Сложный
путь синтеза холестерола можно разделить
на 3 этапа Первый этап заканчивается
образованием мевалоната (мевалоновой
кислоты). Две молекулы ацетил-КоА
конденсируются ферментом тиолазой с
образованием ацетоацетил-КоА. Фермент
щдроксиметилглутарил-КоА-синтаза
присоединяет третий ацетильный остаток
с образованием ГМГ-КоА
(3-гидрокси-3-метилглутарил-КоА). Эта
последовательность реакций сходна с
начальными стадиями синтеза кетоновых
тел (см. рис. 8-33). Однако реакции синтеза
кетоновых тел происходят в митохондриях
печени, а реакции синтеза холестерола
— в цитозоле клеток. Следующая реакция,
катализируемая ГМГ-КоА-редуктазой,
является регуляторной в метаболическом
пути синтеза холестерола. В этой реакции
происходит восстановление ГМГ-КоА до
мевалоната с использованием 2 молекул
NADPH. Фермент ГМГ-КоА-редуктаза —
гликопротеин, пронизывающий мембрану
ЭР, активный центр которого выступает
в цитозоль.На втором этапе синтеза
мевалонат превращается в пятиуглеродную
изопреноидную структуру, содержащую
пирофосфат — изопентенилпирофосфат.
Продукт конденсации 2 изопреновых единиц
— геранилпирофосфат. Присоединение ещё
1 изопреновой единицы приводит к
образованию фарнезилпирофосфата —
соединения, состоящего из 15 углеродных
атомов. Две молекулы фарнезилпирофосфата
конденсируются с образованием сквалена
— углеводорода линейной структуры,
состоящего из 30 углеродных атомов.На
третьем этапе синтеза холестерола
сквален через стадию образования
эпоксида ферментом циклазой превращается
в молекулу ланостерола, содержащую 4
конденсированных цикла и 30 атомов
углерода. Далее происходит 20 последовательных
реакций, превращающих ланостерол в
холестерол. На последних этапах синтеза
от ланостерола отделяется 3 атома
углерода, поэтому холестерол содержит
27 углеродных атомов. У холестерола
имеется насыщенная разветвлённая
боковая цепь из 8 углеродных атомов в
положении 17, двойная связь в кольце В
между атомами углерода в положениях 5
и 6, а также гидроксильная группа в
положении 3. В организме человека
изопентенилпирофосфат также служит
предшественником убихинона (KoQ) и
долихола, участвующего в синтезе
гликопротеинов. Регуляция ключевого
фермента синтеза холестерола
(ГМГ-КоА-редуктазы) происходит разными
способами.
Фосфорилирование/дефосфорилирование
ГМГ-КоА-редуктазы (рис. 8-67). При увеличении
соотношения инеулин/глюкагон этот
фермент дефосфорилируется и переходит
в активное состояние. Действие инсулина
осуществляется через 2 фермента:
фосфатазу
киназы ГМГ-КоА-редуктазы, которая
превращает киназу в неактивное
дефосфорилированное состояние;фосфатазу
ГМГ-КоА-редуктазы путём превращения
её в дефосфорилированное активное
состояние. Результатом этих реакций
служит образование дефосфорилированной
активной формы ГМГ-КоА-редуктазы.
Следовательно,
в абсорбтивный период синтез холестерола
увеличивается. В этот период увеличивается
и доступность исходного субстрата для
синтеза холестерола — ацетил-КоА (в
результате приёма пищи, содержащей
углеводы и жиры, так как ацетил-КоА
образуется при распаде глюкозы и жирных
кислот). В постабсорбтивном состоянии
глюкагон через протеинкиназу А стимулирует
фосфорилирование ГМГ-КоА-редуктазы,
переводя её в неактивное состояние. Это
действие усиливается тем, что одновременно
глюкагон стимулирует фосфорилирование
и инактивацию фосфатазы ГМГ-КоА-редуктазы
и фосфорилирование киназы ГМГ-КоА-редуктазы,
удерживая, таким образом, ГМГ-КоА-редуктазу
в фосфорилированном неактивном состоянии.
В результате синтез холестерола в
постабсорбтивном периоде и при голодании
ингибируется. Ингибирование синтеза
ГМГ-КоА-редуктазы. Конечный продукт
метаболического пути (холестерол)
снижает скорость транскрипции гена
ГМГ-КоА-редуктазы, подавляя таким образом
собственный синтез. В печени активно
идёт синтез жёлчных кислот из холестерола,
поэтому и жёлчные кислоты (как конечные
продукты синтеза) подавляют активность
гена ГМГ-КоА-редуктазы (рис. 8-67). Так как
молекула ГМГ-КоА-редуктазы существует
около 3 ч после синтеза, то ингибирование
синтеза этого фермента кочечным продуктом
метаболического пути (холестеролом)
является эффективной регуляцией.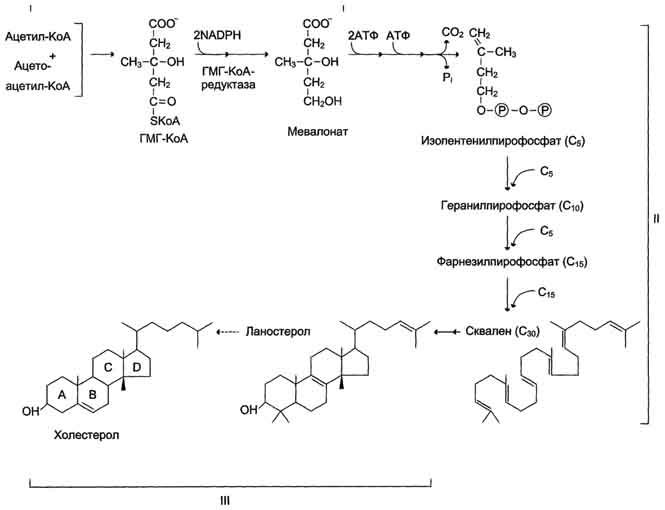 Вопрос
Вопрос
55. ЛПНП и ЛПВП как транспортные формы
холестерина в крови, их роль в обмене
холестерина. Атерогенные и атерогенные
ЛП. Рецепторы ЛПНП.
На ЛППП,
оставшиеся в крови, продолжает действовать
ЛП-липаза, и они превращаются в ЛПНП,
содержащие до 55% холестерола и его
эфиров. Апопротеины Е и С-II реносятся
обратно в ЛПВП. Поэтому основным
апопротеином в ЛПНП служит апоВ-100.
Апопротеин В-100 взаимодействует с
рецепторами ЛПНП и таким образом
определяет дальнейший путь холестерола.
ЛПНП — основная транспортная форма
холестерола, в которой он доставляется
в ткани. Около 70% холестерола и его эфиров
в крови находится в составе ЛПНП. Из
крови ЛПНП поступают в печень (до 75%) и
другие ткани, которые имеют на своей
поверхности рецепторы ЛПНП. Рецептор
ЛПНП — сложный белок, состоящий из 5
доменов и содержащий углеводную часть
рецепторы ЛПНП синтезируются в ЭР и
аппарате Гольджи, а затем экспонируются
на поверхности клетки, в специальных
углублениях, выстланных белком клатрином.
Эти углубления называют окаймлёнными
ямками. Выступающий на поверхность
N-концевой домен рецептора взаимодействует
с белками апоВ-100 и апоЕ; поэтому он может
связывать не только ЛПНП, но и ЛППП,
ЛПОНП, остаточные ХМ, содержащие эти
апопротеины. Клетки тканей содержат
большое количество рецепторов ЛПНП на
своей поверхности: например, на одной
клетке фибробласта имеется от 20 000 до
50 000 рецепторов. Из этого следует, что
холестерол поступает в клетки из крови
в основном в составе лпнп. Если количество
холестерола, поступающего в клетку,
превышает её потребность, то синтез
рецепторов ЛПНП подавляется, что
уменьшает поток холестерола из крови
в клетки. При снижении концентрации
свободного холестерола в клетке,
наоборот, активируется синтез
ГМГ-КоА-редуктазы и рецепторов ЛПНП.
В регуляции синтеза рецепторов ЛПНП
участвуют гормоны: инсулин и трийодтиронин
(Т3), полрвые гормоны. Они увеличивают
образование рецепторов ЛПНП, а
глюкокортикоиды(в основном кортизол)
уменьшают. Эффекты инсулина и Т3, вероятно,
могут объяснить механизм гиперхолестеролемии
и увеличение риска атеросклероза при
сахарном диабете или гипотиреозе. ЛПВП
выполняют 2 основные функции: они
поставляют апопротеины другим ЛП в
крови и участвуют в так называемом
«обратном транспорте холестерола».
ЛПВП синтезируются в печени и в небольшом
количестве в тонком кишечнике в виде
«незрелых липопротеинов» —
предшественников ЛПВП. Они имеют
дисковидную форму, небольшой размер и
содержат высокий процент белков и
фосфолипидов. В печени в ЛПВП включаются
апопротеины А, Е, С-II, фермент ЛХАТ. В
крови апоС-II и апоЕ переносятся с ЛПВП
на ХМ и ЛПОНП. Предшественники ЛПВП
пракгически не содержат холестерола и
ТАГ и в крови обогащаются холестеролом,
получая его из других ЛП и мембран
клеток. Для переноса холестерола в
ЛПВП существует сложный механизм. На
поверхности ЛПВП находится фермент
ЛХАТ — лецитишхолестерол-ацилтрансфераза.
Этот фермент превращает холестерол,
имеющий гидроксильную группу, выступающую
на поверхность липопротеинов или мембран
клеток, в эфиры холестерола. Радикал
жирной кислоты переносится от
фосфатидилхолита(лецитина) на гидроксильную
группу холестерола. Реакция активируется
апопротеином A-I, входящим в состав ЛПВП.
Гидрофобная молекула, эфира холестерола
перемещается внутрь ЛПВП. Таким образом,
частицы ЛПВП обогащаются эфирами
холестерола. ЛПВП увеличиваются в
размерах, из дисковидных небольших
частиц превращаются в частицы сферической
формы, которые называют ЛПВП3, или «зрелые
ЛПВП». ЛПВП3 частично обменивают эфиры
холестерола на триацилглицеролы,
содержащиеся в ЛПОНП, ЛППП и ХМ (рис.
8-70). В этом переносе участвует «белок,
переносящий эфиры холестерина» (он
также называется aпoD). Таким образом,
часть эфиров холестерола переносится
на ЛПОНП, ЛППП, а ЛПВП3 за счёт накопления
триацилглицеролов увеличиваются в
размерах и превращаются в ЛПВП2. ЛПОНП
под действием ЛП-липазы превращаются
сначала в ЛППП, а затем в ЛПНП. ЛПНП и
ЛППП захватываются клетками через
рецепторы ЛПНП. Таким образом, холестерол
из всех тканей возвращается в печень в
основном в составе ЛПНП, но в этом
участвуют также ЛППП и ЛПВП2. Практически
весь холестерол, который должен быть
выведен из организма, поступает в печень
и уже из этого органа выделяется в виде
производных с фекалиями. Путь возвращения
холестерола в печень называют «обратным
транспортом» холестерола.
Соседние файлы в предмете Биохимия
- #
- #
- #
- #
- #
- #
Источник
Регуляция синтеза холестерина — Студопедия
Регуляция синтеза холестерина – его ключевого фермента (ГМГ-КоА редуктазы) проходит разными способами.
Фосфорилирование / дефосфорилирование ГМГ-редуктазы. При увеличении соотношения инсулин/глюкагон этот фермент фосфорилируется и переходит в активное состояние. Действие инсулина осуществляется через 2 фермента.
Фосфатазу киназы ГМГ-КоА-редуктазы, которая привращает киназу в неактивное дефосфорилированное состояние:
Фосфотазу ГМГ-КоА-редуктазы путем превращения ее в дефосфорилированное активное состояние. Результатом этих реакций служит образование дефосорилированной активной формы ГМГ-КоА-редуктазы.
Следовательно, в абсорбтивный период холестерин увеличивается. В этот период увеличивается и доступность исходного субстрата для синтеза холестерина – ацетил – КоА (в результате приема пищи, содержащей углеводы и жиры, так как ацетил КоА образуется при распаде глюкозы и жирных кислот).
В постабсорбтивном состоянии глюкагон через протеингеназу А стимулирует фосфорилирование ГМГ – КоА – редуктазы, переводя ее в неактивное состояние. Это действие усиливается тем, что одновременно глюкагон стимулирует фосфорилирование и инактивацию фосфотазы ГМГ-КоА-редуктазы, удерживая, таким образом, ГМГ-КоА-редуктазу в фосфорилированном неактивном состоянии. В результате синтез холестерина в постабсорбтивном периоде и при голодании ингибируются путем эндогенного синтеза. Если же содержание холестерина в пище доводили до 2%, то синтез эндогенного холестерина резко уменьшался. Но полного прекращения синтеза холестерина не происходит.
Степень ингибирования биосинтеза холестерина под действием холестерина, поступающего с пищей у разных людей различен. Это говорит об индивидуальности процессов образования холестерина. Уменьшая интенсивность синтеза холестерина, можно уменьшить его концентрацию в крови.
Если нарушен баланс между поступлением холестерина с пищей и его синтезом в организме с одной стороны и выведением желчных кислот и холестерина – с другой, концентрация холестерина в тканях и крови изменяется. Наиболее серьезные последствия связаны с повышением концентрации холестерина в крови (гиперхолестеринемия) при этом увеличивается вероятность заболевания атеросклерозом и желчно-каменной болезнью.
Семейная гиперхолестеринемия (ГХС) – эта форма встречается значительно чаще – примерно 1 больной на 200 человек. Наследственный дефект при ГХС заключается в нарушении поглощения ЛПНП клетками, а следовательно, и в снижении скорости катаболизма ЛПНП. Вследствие этого в крови повышается концентрация ЛПНП, а также холестерин поскольку его много в ЛПНП. Поэтому при ГХС характерно отложение холестерина в тканях, в частности в коже (ксантомы), в стенках артерий.
Ингибирование синтеза ГМГ-КоА-редуктазы
Конечный продукт метаболического пути холестерина. Снижает скорость транскрипции гена ГМГ-КоА-редуктазы, подавляя такоим образом собственный синтез. В печени активно идет синтез желчных кислот из холестерина, поэтому и желчные кислоты подавляют активность гена ГМГ-КоА-редуктазы. ГМГ-КоА-редуктаза существует после синтеза около 3, то ингибирование синтеза этого фермента холестерина является эффективной регуляцией.
Источник
биохимия, обмен и его регуляция
В организме человека происходит ежеминутно много биохимических реакций и обменных процессов. Метаболизм не останавливается ни на секунду.
Многие биохимические процессы возглавляют клетки печени, и несут за них ответственность. Основная функция клеток печени — это синтез холестерола.
От данного процесса зависит выработка стероидного типа половых гормонов, преобразование солнечной энергии в витамин Д, и транспортировка питания во все клетки организма при помощи холестериновых молекул.
В чем суть процесса выработки молекул холестерина?
Многие продукты питания наполняют организм холестерином — это продукты животного происхождения, а также транс-жиры, которые находятся в большом количестве в полуфабрикатах промышленного производства, а также в продуктах быстрого питания (фаст-фудах).
Если употреблять такие продукты безмерно, тогда концентрация в составе крови молекул холестерина станет высокой и придётся прибегать к медикаментозному решению гиперхолестеринемии.
Холестерин, который попадает в организм с продуктами питания, имеет низкую молекулярную плотность, что и привод к оседанию такого холестерина на внутренних оболочках сосудов, что провоцирует развитие холестериновой бляшки и патологии атеросклероз.
Повышение индекса холестерина в составе крови происходит не только по причине попадания его извне, но и от нарушения в процессе синтезирования молекул липопротеидов клетками печени.
Синтез холестеринак содержанию ↑
Синтез холестерола в печени
Синтез холестерола в составе организма составляет приблизительно 0,50 -0,80 грамм в сутки.
Распределяется синтез молекул холестерина в организме:
- 50,0% вырабатывается клетками печени;
- 15,0% — 20,0% — отделами тонкого кишечника;
- 10,0% — синтезируется корой надпочечников и клетками кожных покровов.
Все клетки в человеческом организме имеют способность к синтезу липопротеидов.
С продуктами питания поступает в организм молекулы холестерина до 20,0% от всего общего объема — приблизительно 0,40 грамм в сутки.
Выводится за пределы организма липопротеиды при помощи жёлчной кислоты, и в сутки утилизация молекул холестерола жёлчью — не больше, чем 1,0 грамм.
к содержанию ↑
Биосинтез липопротеидов в составе организма
Биосинтез молекул липидов происходит в эндоплазматическом отделе — ретикулуме. Основой для всех атомов молекул углерода является вещество ацетил-SКоА, которое поступает в эндоплазму из митохондрий в молекулах цитрата.
При биосинтезе молекул липопротеидов принимают участие 18 молекул АТФ, а также участниками синтеза становятся 13 молекул НАДФН.
Процесс образования холестерина проходит не менее 30 этапов и реакций в организме.
Поэтапный синтез липопротеидов можно распределить по группам:
вставить активная подсказка — уровень сахара
- Синтез мевалоновой кислоты происходит в процессе кетогенеза первых двух реакций, а после третьего этапа идёт реакция 3-гидрокси-3-метилглутарил-SКоА с молекулой ГМГ-SКоА-редуктазы. От этой реакции и происходит синтезирование мевалонат. Для этой реакции необходимо достаточного количества глюкозы в составе крови. Восполнить ее можно при помощи сладких продуктов и злаковых культур;
- Синтез изопентенилдифосфата происходит после присоединения фосфата к молекулам мевалоновой кислоты и их дегидрации;
- Синтез фарнезилдифосфат происходит после соединения трёх молекул изопентенилдифосфата;
- Синтезирование сквалена — это связывание 2-х молекул фарнезилдифосфата;
- Происходит реакция перехода сквалена в молекулу ланостерола;
- После удаления ненужных метильных групп и происходит преобразование холестерола.
к содержанию ↑
Регуляция процесса синтеза липопротеидов
Регуляторным элементом в процессе синтеза является фермент гидроксиметилглутарил-SКоА-редуктазы. Способность данного фермента изменять активность более чем в 100 раз.
Регуляция активности фермента происходи по нескольким принципам:
- Регуляция синтезирования на метаболическом уровне. Данный принцип работает «от обратного», фермент ингибируется холестеролом, что дает возможность поддерживать постоянное внутриклеточное его содержание;
- Ковалентная гормональная регуляция.
Регуляция на гормональном уровне происходит по таким этапам:
- Повышение содержания в организме гормона инсулина, активирует протеин-фосфатазу, что провоцирует увеличение активности главного фермента ГМГ-SКоА-редуктазы;
- Гормон глюкагон и гормон адреналин имеют свойства активировать элемент протеинкиназы А, которая фосфорилирует фермент ГМГ-SКоА-редуктазы и снижает их активность;
- Активность синтезирования холестерола зависит от концентрации в составе крови специального белка-транспортировщика, что своевременно связывает промежуточные реакции метаболитов.
Регуляция активности гидроксиметилглутарил-S-КоА-редуктазык содержанию ↑
Использование холестерина организмом
Синтезируемый в клетках печени холестерин, необходим организму для различных жизненно важных процессов:
- Находясь в каждой мембране клеток, молекулы холестерина укрепляют их и делают эластичными;
- При помощи липопротеинов, сосудистые оболочки увеличивают свою проницаемость, что защищает их от внешнего воздействия;
- Без помощи липопротеидов надпочечники не производят выработку стероидного типа половых гормонов;
- При помощи липидов происходит продуцирование желчной кислоты и предотвращает желчный пузырь от камнеобразования в нем;
- Липопротеиды связывают между собой клетки нейронов в спинном и в головном мозге;
- При помощи липопротеидов происходит укрепление оболочки нервных волокон;
- С помощью холестерола происходит выработка витамина Д, что помогает усваиваться кальцию и предотвращает разрушение костных тканей.
Холестерол помогает производить надпочечниками синтез таких групп гормонов:
- Группа кортикостероидов;
- Группа гормонов глюкокортикоидов;
- Группа минералокортикоидов.
Холестерол помогает производить надпочечниками синтез групп гормонов
Данные гормоны обеспечивают процессы гормонального регулирования репродуктивных органов человека.
Молекулы холестерина после синтеза в клетках печени попадают в эндокринный орган надпочечники и способствуют выработке гормонов и поддержанию баланса в гормональной сфере.
к содержанию ↑
Выработка молекул витамина Д происходит от солнечного света, который проникает в холестерин, находящийся под кожей. В этот момент и происходит синтез витамина Д, который очень важен для усваивания организмом минералов кальция.
Все виды липопротеидов, после синтеза транспортируются по организму системой кровотока.
Витамин Д может преобразовываться только липопротеидами высокой молекулярной плотности, а низкомолекулярные липиды становятся причиной развития патологии атеросклероз, потому что имеют свойства оседать на внутренних оболочках артерий в виде холестериновых бляшек, которые разрастаясь и провоцируют данную патологию.
Иногда холестериновые бляшки можно наблюдать у человека под кожными покровами на кистях рук.
Метаболизм витамина Дк содержанию ↑
Нарушения в процессе синтеза липопротеидов
Во многих обменных процессах в организме может произойти сбой и нарушение. Такие нарушения могут произойти и в липидном обмене. Причин достаточно много и они имеют экзогенную и эндогенную этиологию.
К эндогенным причинам нарушения синтеза липопротеидов относятся:
- Возраст человека. После 40 лет в организме человека происходит затухание выработки половых гормонов и нарушается гормональный фон, а также к 45 — 50 годам происходит замедление всех обменных процессов, что может внести разлад и в липидный обмен;
- Половой признак — мужчины больше подвержены накапливанию холестерина, чем женщины. Женщины до климактерического периода и менопаузы, защищены выработкой половых гормонов, от накопления липопротеинов;
- Генетическая наследственная предрасположенность. Развитие семейной гиперхолестеремии.
К экзогенным причинам липидного сбоя относятся факторы, что зависят от стиля жизни больного, а также сопутствующие патологии, способствующие нарушению в синтезе молекул холестерина:
- Никотиновая зависимость;
- Хроническая алкогольная зависимость;
- Неправильное питание может привести к повышенному содержанию в организме холестерина и накопления его не только в составе крови;
- Малоподвижный образ жизни становится причиной замедленных обменных процессов и синтеза липопротеидов;
- Гипертоническая болезнь — высокое давление в системе кровотока дает предпосылки тому, чтобы сосудистые оболочки пропитывались липидными жирами, которые впоследствии формируют холестериновую бляшку;
- Заболевание дислипидемия — это нарушение в липидном обмене. При патологии происходит дисбаланс между липопротеидами ВП, липидами НП, а также уровень содержания в составе крови триглицеридов;
- Патология ожирение;
- Сахарный диабет. При гипергликемии нарушается метаболизм и обмен липидами.
Патология ожирениек содержанию ↑
Дефицит в организме молекул полезного холестерола
Существуют патологии, которые снижают концентрацию высокомолекулярного холестерина в составе крови по причине снижения синтеза молекул ЛПВП.
Это может привести к патологиям в щитовидной железе, может значительно повлиять на уровень сахара в составе крови и спровоцировать сахарный диабет, а также стать причиной многих заболеваний системы кровотока и сердечного органа.
Последствиями низкой концентрации высокомолекулярного холестерина могут быть:
- Патология рахит, которая развивается в детском возрасте по причине сниженного синтеза витамина Д и не усвояемости молекул кальция;
- Раннее старение клеток организма. Без своевременной подачи холестерина в клеточные мембраны, происходит их разрушение и запускается процесс старения;
- Резкое снижение массы тела, которое происходит от недостаточного синтеза молекул холестерина, и нарушения в липидном обмене;
- Болезненность в мышечных тканях от недостатка в клетках мышц липидов;
- Боли в сердечном органе, что могут спровоцировать сердечный приступ.
Откорректировать индекс высокомолекулярного холестерина можно при помощи диетического питания, в состав которого входит морская рыба, различные растительные масла, а также кисломолочная продукция.
И не стоит забывать о свежих фруктах, зелени и овощах — они должны преобладать в рационе диеты.
к содержанию ↑
Фотогалерея: Недостаток полезного холестерина
к содержанию ↑
Превышение нормы липопротеидов
Если в человеческом организме происходит усиленный синтез высокомолекулярного липопротеида, тогда это также может негативно сказаться на здоровье.
Повышенная концентрация грозит развитием таких патологий:
- Развивается цирроз клеток печени;
- Риск развития гепатита;
- Увеличение массы тела;
- Нарушение во всем обмене липидов;
- Процессы воспаления в системе кровотока, а также в печеночном органе.
Также от переизбытка молекул холестерина ЛПВП и ЛПНП происходит оседание молекул на стенках артериальных оболочек, образуя холестериновые бляшки, которые влияют на движение крови в русле, по причине сужения артериального просвета.
Также от нарушения в синтезе холестерола, происходит неправильная работа желчного органа, что приводит к камнеобразованию в нем.
к содержанию ↑
Видео: Холестерин — бомба замедленного действия!
к содержанию ↑
Заключение
Синтез холестерола в печени — это очень сложный процесс, который происходит в организме постоянно и не прерываясь.
Если происходят нарушения в липидном обмене, или в процессе синтезирования липопротеидов, тогда существует высокий риск развития патологии атеросклероз, которая прогрессируя, приводит к более тяжелым заболеваниям, что могут спровоцировать преждевременную смерть.
Нарушения в липидном обмене, также влияют на работу сердечного органа и на функциональность клеток печени.
Постоянная корректировка холестерина при помощи диеты, позволит поддерживать липидный обмен в пределах нормы.
Загрузка…
Источник