Что такое сродство гемоглобина к кислороду
Физиология и патофизиология газообмена
Транспорт О2 из атмосферы до периферических тканей
организма представляет собой многоступенчатый процесс.
После того как воздух попадает в альвеолы легких, следующим этапом
газообмена является диффузия кислорода из альвеол в кровь легочных
капилляров и диффузия СО2 из крови легочных капилляров
в альвеолы. Диффузия представляет собой простое движение молекул через
респираторную мембрану из области более высокого давления в область
более низкого.
Таким образом, парциальное давление О2 (PO2)
является одним из основных факторов, определяющих его транспорт в
организме, причем не только в легких, но и во всем теле вплоть до
периферических тканей.
Помимо градиента давления скорость диффузии определяется 1) растворимостью
газа в жидкости; 2) площадью поверхности, через которую протекает
диффузия; 3) расстоянием, которое газ должен пройти при диффузии;
4) молекулярным весом газа; 5) температурой жидкости. Поскольку в
живом организме температура постоянна, она обычно не учитывается.
Применительно к альвеолокапиллярной мембране скорость диффузии газа
будет зависеть от:
- толщины мембраны;
- площади поверхности мембраны;
- диффузионного коэффициента газа в мембране;
- градиента давления газа по обе стороны мембраны.
По мере транспорта О2 от легких к периферическим
тканям его парциальное давление снижается. Если в атмосферном воздухе
при нормальном атмосферном давлении парциальное давление О2
составляет 159 мм рт. ст., то в периферических тканях, в зависимости
от уровня их обмена, — 35–85 мм рт. ст. Венозная кровь, поступающая
в легкие, имеет РО2 около 40 мм рт. ст.
Уже в альвеолярном воздухе содержание О2 и его парциальное
давление отличаются от атмосферного воздуха (13,6 % и 104 мм рт.
ст. соответственно). Это происходит вследствие нескольких причин:
- альвеолярный воздух лишь частично замещается атмосферным во время
каждого вдоха; - О2 постоянно абсорбируется из альвеолярного воздуха;
- СО2 постоянно диффундирует из крови легочных капилляров
в альвеолы; - сухой атмосферный воздух, который поступает в дыхательные пути, увлажняется,
насыщаясь водными парами, прежде чем достигает альвеол.
В артериальной крови, покидающей легкие, РО2
уменьшается уже до 95 мм рт. ст. вследствие так называемого венозного
примешивания. Дело в том, что определенное количество венозной крови
(1–2 %) не аэрируется в легких, поступая напрямую в артериальное
русло, что и приводит к некоторому снижению РО2.
Увеличение объема шунтированной крови, которое происходит при гипоксии
может приводить к существенному понижению артериального РО2.
Увеличение толщины альвеолокапиллярной мембраны, затрудняющее диффузию
газов, часто является следствием отека — увеличения количества жидкости
в межклеточном пространстве мембраны. Состояние характеризуется как
интерстициальный отек легких. Кроме того, жидкость может накапливаться
и в альвеолах, так что газам приходится проходить не только через
мембрану, но и через жидкость, что характерно уже для альвеолярного
отека легких. В большинстве случаев имеет место сочетание этих состояний
той или иной степени выраженности. Имеет значение и исходное состояние
легких. При ряде заболеваний, может возникнуть фиброз какого либо
участка легких с утолщением альвеолокапиллярной мембраны. Поскольку
скорость диффузии газов через мембрану обратно пропорциональна ее
толщине, любой фактор, увеличивающий ее толщину более чем в два –
три раза, может существенно нарушать нормальный газообмен.
Из капилляров кровеносного русла О2 диффундирует
в периферические ткани, поскольку парциальное давление О2
в капиллярах выше, чем в тканях. РО2 в интерстициальной
жидкости вне капилляров составляет в среднем 40 мм рт. ст., тогда
как в артериальной крови — 95 мм рт. ст. В венозной крови, покидающей
капилляр, РО2 также составляет около 40 мм рт. ст.
Если скорость тканевого кровотока увеличивается, большие количества
О2 доставляются тканям и тканевое РО2
увеличивается. Увеличение тканевого метаболизма приводит к снижению
РО2 как в периферических тканях, так и в венозной
крови.
Поскольку О2 постоянно используется тканями, внутриклеточное
РО2 всегда остается ниже интерстициального. Со снижением
интерстициального РО2 снижается и РО2
внутри клеток, повышение интерстициального РО2 ведет
к повышению внутриклеточного РО2. Важным общим свойством
живых организмов является большой запас функциональной прочности систем
поддержания жизнедеятельности. Такая особенность характерна и для
процессов газообмена. Для полного обеспечения процессов метаболизма
достаточно РО2 1–5 мм рт. ст., в то время как внутриклеточное
РО2 может варьировать от 5 до 60 мм рт. ст. (в зависимости
от протяженности капиллярного русла). Поэтому организм человека и
может функционировать при весьма существенном снижении РО2
в атмосферном воздухе.
В нормальных условиях около 97 % кислорода от легких к тканям переносится
в химически связанном виде гемоглобином. Лишь 3 % составляет О2
растворенный в плазме крови. Следует учитывать, что способность гемоглобина
связывать О2 является ограниченной. Каждый грамм
гемоглобина может максимально связать 1,34 мл О2.
Эта так называемая константа Гюффнера. Соответственно кислородная
емкость крови (т. е. максимальное общее количество кислорода, которое
может быть перенесено кровью) будет находиться в прямой зависимости
от содержания гемоглобина:
Кислородная емкость крови = [ Hb ]
x
1, 34 мл O2 / 100 мл крови
У здоровых людей с содержанием гемоглобина 150 г/л кислородная емкость
крови составляет 201 мл О2/л крови. Реально переносимое
количество мл О2 обычно меньше.
Ключевым фактором, характеризующим количество кислорода, связанного
с гемоглобином, является насыщение артериальной крови кислородом (сатурация,
SaO2). Оно выражает отношение между количеством кислорода,
связанного с гемоглобином и кислородной емкостью крови:
SaO2 = ( HbO2 / кислородная емкость крови )
x
100 %
Содержание кислорода в крови (контент, СаО2) — это
сумма связанного с гемоглобином и растворенного в плазме О2:
CaO2 = ( 1, 34
x
[ Hb ]
x
SaO2 ) + ( PaO2
x
0, 0031 )
Очевидно, что СаО2 зависит главным образом от фракции
О2, связанной с гемоглобином.
Кровь содержит незначительное количество кислорода, не связанного
с гемоглобином, а растворенного в плазме. Согласно закону Генри, количество
растворенного кислорода пропорционально парциальному давлению О2
и коэффициенту его растворимости, а растворимость О2
в крови очень низка: только 0,0031 мл О2 растворяется
в 0,1 л крови при увеличении давления на 1 мм рт. ст. Таким образом,
при РаО2 равным 100 мм рт. ст., в 100 мл артериальной
крови содержится только 0,31 мл растворенного О2.
Со снижением РаО2 количество растворенного в плазме
О2 станет еще меньше.
Содержание О2 в связи с изменениями РаО2
колеблется незначительно до тех пор, пока устойчиво поддерживается
SaO2. Изменения содержания гемоглобина приводят к
более заметным сдвигам СаО2. Нормальное СаО2
равно 198 мл О2/л крови при условии, что
PaO2 = 100 мм рт. ст.,
содержание гемоглобина 150 г/л, а
SaO2 = 97 %.
Умеренная анемия (например гемоглобин 120 г/л) при поддержании нормального
РаО2 проявляется снижением СаО2
до 160 мл О2/л крови.
Сродство гемоглобина к кислороду возрастает по мере последовательного
связывания молекул О2, что придает кривой диссоциации
оксигемоглобина сигмовидную или S-образную форму. Эта кривая, соотносящая
изменения SaO2 в зависимости от РаО2,
важна для анализа процессов транспорта кислорода к периферическим
тканям:
Верхняя часть кривой (при РаО2 > 60
мм рт. ст.) — относительно плоская. Это приводит к тому, что SaO2,
а следовательно, и СаО2 остаются достаточно постоянными,
несмотря на значительные колебания РаО2. Повышение
СаО2 или транспорта кислорода в этой области кривой
может быть достигнуто только за счет увеличения содержания гемоглобина
(например, при переливании крови) или растворения в плазме крови кислорода
(например при гипербарической оксигенации).
Крутые средняя и нижняя часть кривой иллюстрируют то положение, что,
хотя SaO2 падает (когда РаО2 оказывается
ниже 60 мм рт. ст.), процесс насыщения гемоглобина кислородом продолжается,
поскольку градиент РаО2 между альвеолами и капиллярами
сохраняется. Периферические ткани в этих условиях могут продолжать
извлекать достаточное количество О2, несмотря на
снижение капиллярного РО2.
РаО2, при котором гемоглобин насыщен кислородом на
50 % (при 37 °С и рН 7,4) известно как Р50. Это
общепринятая мера сродства гемоглобина к кислороду. Р50
в крови человека в норме составляет 26,6 мм. рт. ст. Однако оно может
изменяться при различных метаболических и физиологических условиях,
воздействующих на процесс связывания кислорода гемоглобином.
Когда сродство гемоглобина к кислороду падает, О2
с большой легкостью переходит в ткани, и наоборот. Повышение Р50
определяет сдвиг кривой диссоциации оксигемоглобина вправо. Сродство
гемоглобина к кислороду снижается, указывая, что теперь требуется
более высокое РаО2 для поддержания SaO2
на прежнем уровне. Более низкое сродство гемоглобина к кислороду означает
повышенное высвобождение кислорода в тканях, но ухудшение связывания
гемоглобина с О2 в легких.
Сдвиг кривой диссоциации влево и соответствующее снижение Р50
указывает на повышенное сродство гемоглобина к кислороду — улучшение
связывания в легких и ухудшение высвобождения О2
в периферических тканях. На Р50 и положение кривой
диссоциации гемоглобина влияет несколько факторов, в частности, рН
и температура.
Для обеспечения обмена веществ в периферических тканях важное значение
имеет не только общее содержание О2 — ключевой физиологической
переменной является его доставка. Доставка кислорода — это количество
кислорода, транспортируемое к тканям в единицу времени. Чтобы поддержать
аэробный метаболизм и предотвратить накопление лактата, периферические
ткани должны постоянно снабжаться кислородом. Обстоятельства, которые
определяют адекватность кислородного снабжения, разнообразны и включают
состояние покоя, физическую нагрузку, гиперкатаболические состояния
и инфекцию.
Доставка кислорода к периферическим тканям зависит от количества кислорода
в определенном объеме крови и уровня кровотока. Хотя объем кровотока
к отдельным органам различен, в периферических тканях он в целом равен
сердечному выбросу (CB). Системная доставка О2 (DО2)
рассчитывается как
DO2 = СВ (л/мин) x CaO2 (мл/л).
Нередко DО2 рассчитывают с учетом площади тела. То
есть на самом деле рассчитывают индекс DО2. Нормальными
величинами индекса DО2 считают 520–720 мл·мин-1·м-2.
Доставка кислорода падает при уменьшении сердечного выброса или снижении
объемного содержания кислорода в артериальной крови. Это характерно
для многих критических и терминальных состояний и чаще всего наблюдается
при выраженных проявлениях сердечной недостаточности, повышении внутрилегочного
шунтирования крови, централизации кровообращения, при различных пороках
сердца, кардиохирургических и других длительных и травматичных вмешательствах.
С практической точки зрения следует иметь в виду, что любое нарушение
кровообращения будет ухудшать доставку О2.
Потребление кислорода является заключительным этапом транспорта кислорода
тканям и представляет собой кислородное обеспечение тканевого метаболизма.
В условиях основного обмена взрослый человек потребляет около 250
мл О2 в 1 мин. Однако скорость утилизации О2
различными тканями значительно отличается.
Потребление кислорода тканями (VO2) — интегральный
показатель, учитывающий как циркуляторный компонент транспорта кислорода
(сердечный выброс), так и его гемический компонент (артериовенозное
различие по кислороду,
CaO2 — CvO2).
Его можно определить по формуле:
VO2 = СВ x (CaO2 — CvO2) x 10.
Так же как и DО2, VO2 часто выражают
в виде индекса, то есть в перерасчете на площадь тела. Нормальными
величинами индекса потребления кислорода являются 110–160 мл·мин-1·м-2.
В зависимости от состояния организма (покой, нагрузка или заболевание)
меняется фракционное распределение СВ к органам. Более того, и экстракция
кислорода тканями различных органов неодинакова. Например, миокард
получает лишь малую фракцию СВ, но извлекает почти весь доставляемый
кислород.
Нормальный компенсаторный ответ на снижение кровотока проявляется
в виде увеличения поглощения кислорода, достаточного для поддержания
VO2 на нормальном уровне. Падение сердечного выброса
компенсируется увеличением артериовенозного различия по кислороду,
и VO2 остается неизменным. Снижение насыщения венозной
крови кислородом отражает увеличение экстракции кислорода.
Способность компенсировать снижение кровотока повышением поглощения
кислорода является характерной особенностью микроциркуляторного русла
практически всех органов и тканей за исключением сердца и диафрагмы.
В них высокая экстракция кислорода из капиллярного ложа происходит
уже в норме. Поэтому уровень кислорода в тканях сердца и диафрагмы
весьма чувствителен даже к незначительным изменениям кровотока.
Таким образом, основными факторами, определяющими доставку О2,
являются:
- парциальное давление кислорода в атмосферном воздухе;
- нормальное состояние альвеолокапиллярной мембраны;
- эффективность дыхательной системы;
- достаточное количество гемоглобина крови;
- эффективность системы кровообращения.
Сергей Науменко,
г. Новосибирск, 2018 г.
Источник
Гемоглобин: роль в газообмене и процессе дыхания
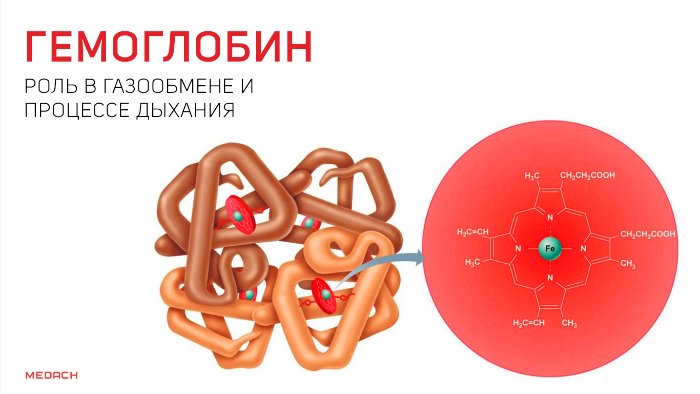
Одним из самых сложных процессов, что происходят в организме человека, несомненно, является дыхание. И сложность эта не только в танце легких, благодаря которому человек получает кислород, но и в процессах, с помощью которых этот кислород проникает дальше, в ткани, где превращается в углекислый газ, что отправляется в обратное путешествие. О данных процессах и пойдет речь далее.
Итак, приступим. Человек делает вдох, иии… Далеко не весь кислород поступает в легкие, а затем и в кровь. Часть вдыхаемого воздуха остается в так называемом мертвом пространстве. Мертвое пространство, в свою очередь, делится на анатомическое (дыхательные пути), в котором остается около 30 % вдыхаемого воздуха, и функциональное (вентилируемые, но по каким-то причинам не перфузируемые альвеолы).
Ухудшение альвеолярного газообмена может происходить при неглубоком и частом дыхании (причиной может стать перелом ребер, паралич дыхательной мускулатуры различного генеза и др.), а также при увеличении мертвого пространства, вызванном разнообразными причинами (нарушение перфузии альвеол в результате воспалительных заболеваний легких, удаление доли или целого легкого и др.), при снижении скорости кровотока по альвеолярным капиллярам (ТЭЛА, инфаркт легкого), при наличии диффузионного барьера (отек легких) и в результате ослабления альвеолярной вентиляции при обтурации просвета бронха. Газообмен между легкими и кровью происходит путем диффузии в соответствии с законом Фика. В легочных капиллярах она происходит за счет разности парциальных давлений в альвеолах и эритроцитах.
В альвеолах парциальное давление кислорода значительно превышает таковое для углекислого газа и составляет примерно 13,3 кПа (100 мм рт. ст.) и 5,3 кПа (40 мм рт. ст.) соответственно. Альвеолы омываются приносимой легочными артериями венозной кровью, в которой соотношение парциальных давлений этих двух газов обратно пропорционально и составляет приблизительно 5,3 кПа (40 мм рт. ст.) для кислорода и 6,1 кПа (46 мм рт. ст.) для углекислого газа. В среднем разница парциальных давлений составляет около 8 кПа (60 мм рт. ст.) для кислорода и около 0,8 кПа для углекислого газа.
Как уже было сказано выше, кислород путем диффузии проникает в кровь легочных капилляров. Диффузионное расстояние для кислорода при этом составляет 1–2 мкм, то есть именно на такое расстояние он проникает внутрь капилляра. Обмен крови в легочном капилляре происходит примерно за 0,75 секунды, но этого времени хватает на то, чтобы парциальные давления в альвеолах и в крови пришли в равновесие.
Кровь, в которой показатели парциального давления для кислорода и углекислого газа примерно равны таковым в альвеолах, называется артериализированной. Однако за счет наличия в легких артериовенозных шунтов и притока венозной крови из бронхиальных вен такой она остается недолго. В результате парциальное давление кислорода в аорте составляет примерно 12,0 кПа (как уже было сказано выше, парциальное давление в артериализированной крови равно таковому в альвеолах и составляет 13,3 кПа), а давление углекислого газа меняется незначительно и не приводит к затруднению его диффузии из крови в альвеолы.
Но кислород непосредственно в ткани попадает лишь в крайне незначительных количествах: для свободного перемещения по организму ему необходим транспортер. Эту функцию выполняет содержащийся в эритроцитах белок — гемоглобин. Гемоглобин существует в оксигенированной и неоксигенированной формах. В дезокси-гемоглобине железо находится на уровне порфиринового кольца и стабилизируется электростатическими силами, что обеспечивает поддержание всей структуры. Появившись, кислород начинает «тянуть» за железо, которое переносится на проксимальный гистидин на другом конце полипептидной цепи и меняет структуру всего протеина.
В результате гемоглобин переходит в оксигенированную форму, альфа- и бета-цепи при этом поворачиваются относительно друг друга на 15 градусов, облегчая присоединение остальных молекул кислорода. В итоге каждый из четырех содержащихся в нем атомов двухвалентного железа обратимо связывается с молекулой кислорода, что превращает молекулу гемоглобина в оксигемоглобин. По сравнению с миоглобином гемоглобин имеет низкое сродство к кислороду, однако оно не статично. Так, миоглобин может связывать кислород только одним участком, поэтому кривая его связывания — гипербола. Кривая связывания гемоглобина с кислородом имеет S-образную форму, демонстрируя, что при его связывании с первой молекулой кислорода гемоглобин имеет очень низкое сродство к кислороду, но при связывании последующих молекул кислорода сродство остальных его субъединиц к нему значительно увеличивается и в конечном счете повышается примерно в 500 раз.
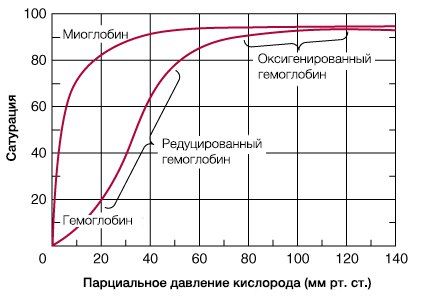
При этом альфа-цепи связывают кислород легче, чем бета-цепи. Этот процесс назван кооперативным взаимодействием. По мере снижения парциального давления кислорода в крови происходит его высвобождение из гемоглобина и поступление в ткани. Например, парциальное давление кислорода в работающих мышцах составляет всего 26 мм рт. ст, и при прохождении эритроцитов через капилляры, кровоснабжающие мышцы, происходит высвобождение и поступление в мышечные клетки примерно ⅓ всего переносимого гемоглобином кислорода. При повышении температуры тела также возрастает потребность в кислороде, что, в свою очередь, стимулирует высвобождение и поступление его в ткани. При снижении температуры, напротив, развивается гипоксия тканей, способствующая компенсаторному увеличению сродства гемоглобина к кислороду.
Гемоглобин также осуществляет перенос от тканей к легким продуктов тканевого дыхания — углекислого газа и ионов водорода. В ходе окислительных процессов в клетке выделяется углекислый газ, в результате гидратации которого образуются ионы водорода, что, в свою очередь, приводит к снижению рН. Давно известно, что снижение рН и повышение концентрации углекислого газа в крови оказывает сильное влияние на способность гемоглобина связывать кислород.
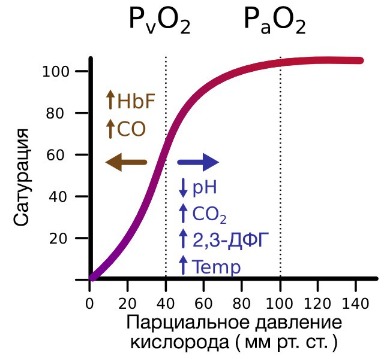
В периферических сосудах показатели рН низкие, и по мере связывания гемоглобина с ионами водорода и углекислым газом происходит снижение его сродства к кислороду. Это влияние величины рН и концентрации углекислого газа на способность гемоглобина связывать кислород называют эффектом Бора.
Обратная ситуация имеет место в альвеолярных капиллярах, где присоединение кислорода к гемоглобину превращает тот в более сильную кислоту.
При этом сродство гемоглобина к углекислому газу снижается, а повышение кислотности гемоглобина приводит к высвобождению излишка ионов водорода и образованию в крови из бикарбоната угольной кислоты, которая затем распадается на воду и углекислый газ. В обоих случаях углекислый газ из крови поступает в альвеолы, а затем в атмосферу. Данный процесс назван эффектом Холдейна. Стоит отметить, что важную роль в образовании углекислого газа в эритроцитах играет ион хлора, поступающий в плазму крови в обмен на бикарбонат при участии белка-переносчика АЕ1. Данный процесс в англоязычной литературе получил название «Chloride shift» или «перенос Хамбургера».
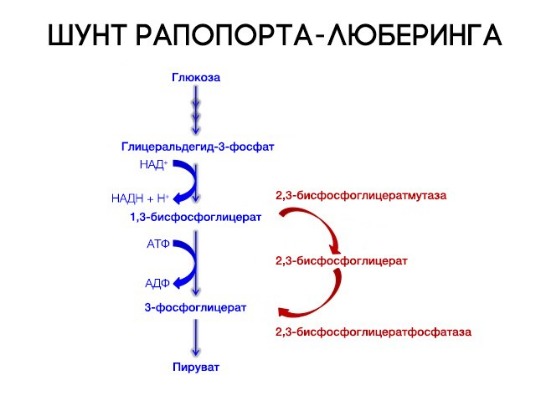
На сродство гемоглобина к кислороду оказывает влияние и присутствующее в эритроцитах вещество, получившее название 2,3-бисфосфоглицерат (БФГ). Его образование — своего рода побочная реакция анаэробного гликолиза, происходящего в эритроцитах в ходе ферментативного превращения глюкозы в пируват под действием фермента бифосфоглицератмутазы. БФГ способен самостоятельно связываться с неоксигенированной формой гемоглобина, образуя солевой мостик между двумя его бета-субъединицами и снижая сродство к кислороду.
При этом гемоглобин способен связать только одну молекулу БФГ, а при присоединении кислорода БФГ вытесняется из полости. В обычных условиях в эритроцитах крови содержится достаточно большое количество БФГ, которое может увеличиваться в условиях гипоксии (например, у дайверов при погружении на глубину), а также при восхождении на большую высоту. В первые часы подъема концентрация БФГ в эритроцитах будет возрастать, а сродство кислороду снижаться. Но на большой высоте парциальное давление будет значительно ниже такового на уровне моря, а значит, оно снизится и в тканях. При этом БФГ будет облегчать передачу кислорода от гемоглобина к тканям.
Некоторые вещества способны прочно связываться с гемоглобином или же вовсе менять его структуру. Одним из них является угарный газ, чье сродство к гемоглобину в 200 раз превышает таковое для кислорода. Отравления угарным газом часто происходят в помещениях с печным отоплением, при пожарах и авариях на производстве. Со временем кислород вытесняет угарный газ из гемоглобина, и в легких случаях пациенты помещаются под наблюдение и получают ингаляции с увлажненным кислородом. Необходимой мерой при тяжелых отравлениях угарным газом является переливание эритроцитарной массы.
К веществам, способным изменять структуру гемоглобина, относятся метгемоглобинобразователи — соединения, способные окислять двухвалентное железо в геме до трехвалентного. К ним относятся нитриты, нитраты, некоторые местные анестетики, аминофенолы, хлораты, примахин и некоторые сульфаниламиды. Состояние, характеризующееся появлением в крови окисленного гемоглобина, называют метгемоглобинемией. При высокой метгемоглобинемии капля крови, помещенная на фильтровальную бумагу, имеет характерный коричневый цвет, а при пропускании кислорода через пробирку с такой кровью ее цвет не меняется. Метгемоглобинемия выше 70 % от общего содержания гемоглобина часто приводит к гибели пациента еще до момента постановки диагноза.
Источники:
- Harrison’s hematology and oncology Longo, Dan L (Dan Louis), Third edition. New York : McGraw-Hill Education Medical, 2017.
- Наглядная физиология, С. Зильбернагль, А. Деспопулос, 2013.
- Ленинджер А. Основы биохимии: В 3-х т. Т. 1. /Д. Нельсон, М. Кокс ; Пер. с англ.-М.: БИНОМ: Лаборатория знаний, 2011.- 694 с.
Нашли опечатку? Выделите фрагмент и нажмите Ctrl+Enter.
Источник