Холестерин и желчные кислоты биологическая роль
Содержание статьи
Без желчи липиды не переварятся
Желчь представляет собой сложную жидкость со щелочной реакцией. В ней выделяют сухой остаток – около 3% и воду – 97%. В сухом остатке обнаруживается две группы веществ:
- попавшие сюда путем фильтрации из крови натрий, калий, бикарбонат-ионы (HCO3¯), креатинин, холестерол (ХС), фосфатидилхолин (ФХ),
- активно секретируемые гепатоцитами билирубин и желчные кислоты.
В норме между основными компонентами желчи Желчные кислоты : Фосфатидилхолин : Холестерин выдерживается соотношение равное 65 : 12 : 5.
В сутки образуется около 10 мл желчи на кг массы тела, таким образом, у взрослого человека это составляет 500-700 мл. Желчеобразование идет непрерывно, хотя интенсивность на протяжении суток резко колеблется.
Роль желчи
1. Наряду с панкреатическим соком нейтрализация кислого химуса, поступающего из желудка. При этом ионы HCO3¯ взаимодействуют с НСl, выделяется углекислый газ и происходит разрыхление химуса, что облегчает переваривание.
2. Обеспечивает переваривание жиров:
- эмульгирование для последующего воздействия липазой, необходима комбинация [желчные кислоты+жирные кислоты+моноацилглицеролы],
- уменьшает поверхностное натяжение, что препятствует сливанию капель жира,
- образование мицелл, способных всасываться.
3. Благодаря п.п.1 и 2 обеспечивает всасывание жирорастворимых витаминов (витамин A, витамин D, витамин K, витамин E).
4. Усиливает перистальтику кишечника.
5. Экскреция избытка ХС, желчных пигментов, креатинина, металлов Zn, Cu, Hg, лекарств. Для холестерина желчь – единственный путь выведения, с ней может выводиться 1-2 г/сут.
Формирование желчи (холерез) идет непрерывно, не прекращаясь даже при голодании. Усиление холереза происходит под влиянием n.vagus и при приеме мясной и жирной пищи. Снижение – под влиянием симпатической нервной системы и повышения гидростатического давления в желчных путях.
Желчевыделение (холекинез) обеспечено низким давлением в двенадцатиперстной кишке, усиливается под влиянием n.vagus и ослабляется симпатической нервной системой. Сокращение желчного пузыря стимулируется бомбезином, секретином, инсулином и холецистокинин—панкреозимином. Расслабление вызывают глюкагон и кальцитонин.
Синтез желчных кислот
Образование желчных кислот идет в эндоплазматическом ретикулуме при участии цитохрома Р450, кислорода, НАДФН и аскорбиновой кислоты. 75% холестерина, образуемого в печени, участвует в синтезе желчных кислот.
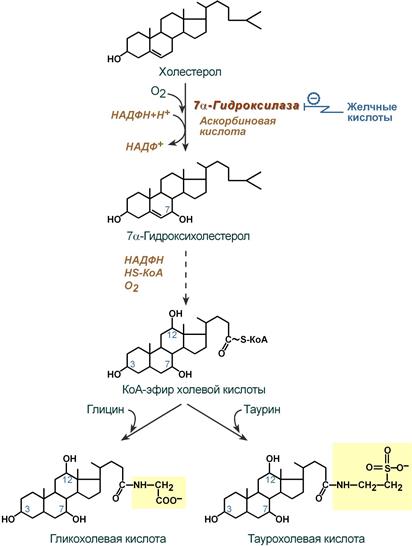
Реакции синтеза желчных кислот на примере холевой кислоты
В печени синтезируются первичные желчные кислоты:
- холевая (3α, 7β, 12α, гидроксилирована по С3, С7, С12),
- хенодезоксихолевая (3α, 7α, гидроксилирована по С3, С7).
Затем они образуют парные желчные киcлоты – конъюгаты с глицином (гликопроизводные) и с таурином (тауропроизводные), в соотношении 3:1 соответственно.
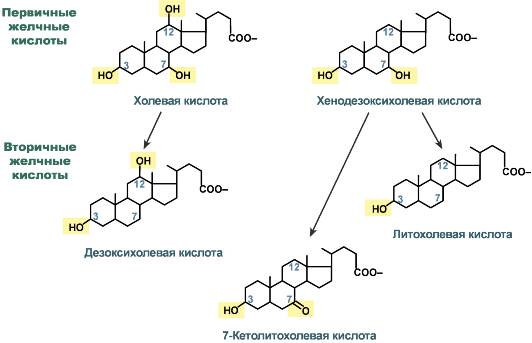
Строение желчных кислот
В кишечнике под действием микрофлоры эти желчные кислоты теряют OH-группу при С7 и превращаются во вторичные желчные кислоты:
- холевая в дезоксихолевую (3α, 12α, гидроксилирована по С3 и С12),
- хенодезоксихолевая в литохолевую (3α, гидроксилирована только по С3) и 7-кетолитохолевую (7α-ОН-группа преобразуется в кетогруппу) кислоты.
Также выделяют третичные желчные кислоты. К ним относятся
- образованная из литохолевой кислоты (3α) – сульфолитохолевая (сульфонирование по C3),
- образованная из 7-кетолитохолевой кислоты (3α, 7-кето) при восстановлении 7-кетогруппы до ОН-группы – урсодезоксихолевая (3α, 7β).
Урсодезоксихолевая кислота является действующим компонентом лекарственного препарата «Урсосан» и используется в терапии заболеваний печени как гепатопротекторное средство. Также она оказывает желчегонное, холелитолитическое, гиполипидемическое, гипохолестеринемическое и иммуномодулирующее действие.
Регуляция синтеза
Регуляторным ферментом является 7α-гидроксилаза. Имеются два способа регуляции
- Коваалентная модификация путем фосфорилирования-дефосфорилирования. Глюкагон, по аденилатциклазному механизму, ведет к фосфорилированию и активации фермента. инсулин оказывает противоположный эффект.
- Генетическая регуляция, т.е. изменение количества фермента:
- она может быть гормональной: тиреоидные гормоны индуцируют транскрипцию и повышают синтез фермента, эстрогены репрессируют и подавляют синтез этого фермента.
- при участии метаболитов: холестерол увеличивает, а желчные кислоты снижают синтез фермента.
Кишечно-печеночная циркуляция
Циркуляция желчных кислот заключается в их непрерывном движении из гепатоцитов в просвет кишечника и реабсорбция большей части желчных кислот в подвздошной кишке, что сберегает ресурсы холестерола. В сутки происходит 6-10 таких циклов. Таким образом, небольшое количество желчных кислот (всего 3-5 г) обеспечивает переваривание липидов, поступающих в течение суток. Потери в размере около 0,5 г/сут соответствуют суточному синтезу холестерола de novo.
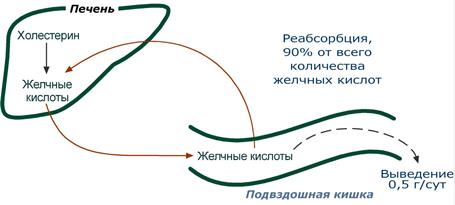
Кишечно-печеночная рециркуляция желчных кислот
Источник
ЖЁЛЧНЫЕ КИСЛОТЫ
ЖЁЛЧНЫЕ КИСЛОТЫ (син. холевые кислоты) — органические кислоты, являющиеся специфическими компонентами желчи и играющие важную роль в переваривании и всасывании жиров, а также в некоторых других процессах, происходящих в желудочно-кишечном тракте, в том числе в переносе липидов в водной среде. Ж. к. представляют собой также конечный продукт обмена холестерина (см.), который выводится из организма в основном в виде Ж. к.
По своей хим. природе Ж. к. являются производными холановой к-ты (C23H39COOH), к кольцевой структуре к-рой присоединены одна, две или три гидроксильные группы. Боковая цепь Ж. к., так же как и в молекуле холановой к-ты, включает 5 углеродных атомов с COOH-группой на конце.
В желчи человека содержатся: холевая (3-альфа,7-альфа,12-альфа-триокси-5-бета-холановая) к-та:

хенодезоксихолевая (антроподезоксихолевая) (3-альфа,7-альфа-диокси-5-бета-холановая) к-та:

и дезоксихолевая (3-альфа,12-альфа-диокси-5-бета-холановая) к-та:

кроме того, в малых количествах или в виде следов содержатся литохолевая (З-альфа-моноокси-5-бета-холановая), а также аллохолевая и урсодезоксихолевая к-ты — стереоизомеры холевой и хенодезоксихолевой к-т. Все Ж. к. присутствуют в желчи (см.) в конъюгированном виде. Часть их конъюгирована с глицином (гликоколом) в гликохолевую или гликохенодезоксихолевую к-ту, а часть — с таурином в таурохолевую:

или таурохенодезоксихолевую к-ту. В печеночной желчи Ж. к. диссоциируют и находятся в форме желчнокислых солей натрия и калия (холатов и дезоксихолатов Na и К), что объясняется щелочными значениями pH желчи (7,5—8,5).
Из всех Ж. к. только холевая и хенодезоксихолевая к-ты первично образуются в печени (их называют первичными), тогда как другие образуются в кишечнике под влиянием ферментов кишечной микрофлоры и носят название вторичных. Они всасываются в кровь и затем вновь секретируются печенью в составе желчи.
У безмикробных животных, выращенных в стерильных условиях, в желчи присутствуют лишь холевая и хенодезоксихолевая к-ты, а дезоксихолевая и литохолевая отсутствуют и появляются в желчи только с внесением в кишечник микроорганизмов. Это подтверждает вторичное образование этих Ж. к. в кишечнике под влиянием микрофлоры из холевой и хенодезоксихолевой к-т соответственно.
Первичные Ж. к. образуются в печени из холестерина.

Этот процесс довольно сложен, т. к. Ж. к. отличаются от холестерина по стереохим. конфигурации двух участков молекулы. Гидроксильная группа у 3-го C-атома в молекуле Ж. к. находится в альфа-положении, a в молекуле холестерина — в бета-положении. Водород у 3-го C-атома Ж. к. находится в p-положении, что соответствует транс-конфигурации колец А и В, а в холестерине — в a-положении (цис-конфигурация колец А и В). Кроме того, Ж. к. содержат большее количество гидроксильных групп, более короткую боковую цепь, которая характеризуется наличием карбоксильной группы.
Процесс превращения холестерина в холевую к-ту начинается с гидроксилирования холестерина в 7альфа-положении, т. е. с включения гидроксильной группы в положение 7, затем следует окисление OH-группы у 3-го С-атома в кетогруппу, перемещение двойной связи от 5-го С-атома к 4-му C-атому, гидроксилирование в 12-альфа-положении и т. д. Все эти реакции катализируются микросомальными ферментами печени в присутствии НАД•H или НАДФ•Н. Окисление боковой цепи в молекуле холестерина осуществляется при участии ряда дегидрогеназ в присутствии АТФ, КоА и ионов Mg2+. Процесс идет через стадию образования 3-альфа, 7-альфа, 12-альфа-триоксикопростановой к-ты, к-рая затем подвергается бета-окислению. В завершающей стадии отделяется трехуглеродный фрагмент, представляющий собой пропионил-КоА, и боковая цепь молекулы, т. о., укорачивается. Последовательность этих реакций в некоторых звеньях может меняться. Напр., образование кетогруппы в 3-бета-положении может происходить не до, а после гидроксилирования в 12-альфа-положении. Однако это не изменяет основного направления процесса.
Процесс образования хенодезоксихолевой к-ты из холестерина имеет некоторые особенности. В частности, окисление боковой цепи с образованием гидроксила у 26-го С-атома может начинаться на каждой стадии процесса, причем гидроксилированный продукт участвует далее в реакциях в обычной последовательности. Возможно, что раннее присоединение OH-группы к 26-му C-атому по сравнению с обычным течением процесса является важным фактором в регуляции синтеза хенодезоксихолевой к-ты. Установлено, что эта к-та не является предшественником холевой и не превращается в нее; равным образом и холевая к-та в организме человека и животных не превращается в хенодезоксихолевую.
Конъюгирование Ж. к. протекает в две стадии. Первая стадия заключается в образовании ацил-КоА, т. е. КоА-эфиров Ж. к. Для первичных Ж. к. эта стадия осуществляется уже на конечном этапе их образования. Вторая стадия конъюгирования Ж. к.— собственно Конъюгирование — состоит в соединении посредством амидной связи молекулы Ж. к. с глицином или таурином. Этот процесс катализируется лизосомальной ацилтрансферазой.
В желчи человека основные Ж. к.— холевая, хенодезоксихолевая и дезоксихолевая — находятся в количественном соотношении 1:1:0,6; глициновые и тауриновые конъюгаты этих к-т — в соотношении 3:1. Соотношение между двумя этими конъюгатами меняется в зависимости от характера пищи: в случае преобладания в ней углеводов увеличивается относительное содержание глициновых конъюгатов, а при высокобелковой диете — тауриновых конъюгатов. Кортикостероидные гормоны повышают относительное содержание тауриновых конъюгатов в желчи. Наоборот, при заболеваниях, сопровождающихся белковой недостаточностью, увеличивается доля глициновых конъюгатов.
Отношение глицин-конъюгированных к таурин-конъюгированным Ж. к. у человека изменяется под влиянием тиреоидного гормона, увеличиваясь при гипотиреоидной состоянии. Кроме того, у больных с гипотиреоидизмом холевая к-та имеет большее время полусуществования и медленнее обменивается, чем у больных с гипертиреоидизмом, что сопровождается повышением содержания холестерина в крови у больных с пониженной функцией щитовидной железы.
У животных и человека при кастрации повышается содержание холестерина в крови. В эксперименте снижение концентрации холестерина в сыворотке крови и увеличение образования Ж. к. наблюдали при введении эстрогена. Тем не менее действие гормонов на биосинтез Ж. к. изучено еще недостаточно.
В желчи различных животных состав Ж. к. сильно варьирует. У многих из них обнаруживаются Ж. к., отсутствующие у человека. Так, у некоторых амфибий главным компонентом желчи является кипринол — желчный спирт, который, в отличие от холевой к-ты, имеет более длинную боковую цепь с двумя гидроксильными группами у 26-го и 27-го С-атомов. Этот спирт конъюгируется преимущественно с сульфатом. У других амфибий преобладает желчный спирт буфол, имеющий ОН-группы у 25-го и 26-го C-атомов. У свиньи в желчи присутствует гиохолевая к-та с OH-группойв положении 6-го С-атома (3-альфа,6-альфа,7-альфа-триоксихолановая к-та). У крыс и мышей имеются альфа- и бета-марихолевые к-ты — стереоизомеры гиохолевой к-ты. У животных, питающихся растительной пищей, в желчи преобладает хенодезоксихолевая к-та. Напр., у морской свинки она является единственной из основных Ж. к. Холевая к-та, напротив, более характерна для плотоядных животных.
Одна из главных функций Ж. к.— перенос липидов в водной среде — связана с их детергентными свойствами, т. е. с их способностью растворять липиды путем образования мицеллярного раствора. Эти свойства Ж. к. проявляются уже в ткани печени, где при их участии из ряда компонентов желчи образуются (или окончательно формируются) мицеллы, получившие название липидного комплекса желчи. Благодаря включению в этот комплекс секретируемые печенью липиды и некоторые другие плохо растворимые в воде вещества переносятся в кишечник в виде гомогенного р-ра в составе желчи.
В кишечнике соли Ж. к. участвуют в эмульгировании жира. Они входят в состав эмульгирующей системы, включающей в себя насыщенный моноглицерид, ненасыщенную жирную к-ту и соли Ж. к. При этом они играют роль стабилизаторов жировой эмульсии. Ж. к. выполняют также важную роль в качестве своеобразного активатора панкреатической липазы (см.). Их активирующее влияние выражается в смещении оптимума действия липазы, который в присутствии Ж. к. перемещается с pH 8,0 до pH 6,0, т. е. до той величины pH, к-рая более постоянно поддерживается в двенадцатиперстной кишке в ходе переваривания жирной пищи.
После расщепления жира липазой продукты этого расщепления — моноглицериды и жирные кислоты (см.) образуют мицеллярный р-р. Решающую роль в этом процессе играют соли Ж. к. Благодаря их детергент-ному действию в кишечнике образуются устойчивые в водной среде мицеллы (см. Молекула), содержащие продукты расщепления жира, холестерин и часто фосфолипиды. В таком виде эти вещества переносятся с эмульсионных частиц, т. е. с места гидролиза липидов, к всасывающей поверхности кишечного эпителия. В виде мицеллярного р-ра, образующегося при участии солей Ж. к., переносятся в жел.-киш. тракте и жирорастворимые витамины. Выключение Ж. к. из пищеварительных процессов, напр, при экспериментальном отведении желчи из кишечника, приводит к снижению всасывания жира в жел.-киш. тракте на 50% и к нарушению всасывания жирорастворимых витаминов вплоть до развития явлений витаминной недостаточности, напр, недостаточности витамина К. Помимо этого, Ж. к. оказывают стимулирующее действие на рост и функции нормальной кишечной микрофлоры: при прекращении поступления желчи в кишечник жизнедеятельность микрофлоры претерпевает существенные изменения.
Выполнив свою физиол, роль в кишечнике, Ж. к. в подавляющем количестве всасываются в кровь, возвращаются к печени и вновь секретируются в составе желчи. Происходит, т. о., постоянная циркуляция Ж. к. между печенью и кишечником. Этот процесс называется печеночно-кишечной (энтерогепатической или портально-билиарной) циркуляцией Ж. к.
Основная масса Ж. к. всасывается в конъюгированном виде в подвздошной кишке. В проксимальной части тонкой кишки нек-рое количество Ж. к. переходит в кровь путем пассивного всасывания.
Исследования, проведенные с помощью меченных 14C Ж. к., показали, что в желчи содержится лишь небольшая часть Ж. к., вновь синтезированных печенью [С. Бергстрем, Даниэльссон (H. Danielsson), 1968]. На их долю приходится только 10—15% от общего количества Ж. к. Основную же массу Ж. к. желчи (85—90%) составляют Ж. к., реабсорбированные в кишечнике и повторно секретируемые в составе желчи, т. е. Ж. к., участвующие в печеночно-кишечной циркуляции. Общий пул Ж. к. у человека в среднем составляет 2,8—3,5 г, причем они совершают 5—6 оборотов за сутки. У разных животных число оборотов, совершаемых Ж. к. за сутки, сильно варьирует: у собаки оно также равно 5—6, а у крысы 10—12.
Часть Ж. к. подвергается в кишечнике деконъюгированию под влиянием нормальной кишечной микрофлоры. При этом нек-рое количество их теряет гидроксильную группу, превращаясь в дезоксихолевую, литохолевую или в другие к-ты. Все они всасываются и после конъюгирования в печени секретируются в составе желчи. Однако 10—15% всех поступивших в кишечник Ж. к. после деконъюгирования подвергается более глубокой деградации. В результате процессов окисления и восстановления, вызываемых ферментами микрофлоры, эти Ж. к. претерпевают различные изменения, сопровождающиеся частичным разрывом их кольцевой структуры. Целый ряд образующихся продуктов затем выделяется с фекалиями.
Биосинтез Ж. к. контролируется по типу отрицательной обратной связи определенным количеством Ж. к., возвращающихся в печень в процессе печеночно-кишечной циркуляции.
Показано, что разные Ж. к. оказывают качественно и количественно различное регулирующее действие. У человека, напр., хенодезоксихолевая к-та тормозит образование холевой к-ты.
Увеличение содержания холестерина в пище приводит к усилению биосинтеза Ж. к.
Разрушение и выброс части Ж. к, представляют важнейший путь экскреции конечных продуктов обмена холестерина. Показано, что у безмикробных животных, лишенных кишечной микрофлоры, сокращается число оборотов, совершаемых Ж. к. между печенью и кишечником, и резко снижается экскреция Ж. к. с фекалиями, что сопровождается повышением содержания холестерина в сыворотке крови.
Т. о., достаточно интенсивная секреция Ж. к. в составе желчи и их превращение в кишечнике под влиянием микрофлоры чрезвычайно важны как для пищеварения, так и для обмена холестерина.
В норме в моче у человека Ж. к. не содержатся, очень небольшие их количества появляются в моче при обтурационной желтухе (ранние стадии) и острых панкреатитах. Ж. к. являются самыми сильными холеретиками, напр, дегидрохолевая кислота (см.). Это свойство Ж. к. используется для введения их в состав желчегонных средств (см.) — дехолина, аллохола и др. Ж. к. стимулируют перистальтику кишечника. Запоры, наблюдаемые у больных с желтухой, могут быть обусловлены дефицитом холатов (солей Ж. к.). Однако одномоментное поступление большого количества конц. желчи в кишечник, а с нею и большого количества Ж. к., наблюдающееся у ряда больных после удаления желчного пузыря, может вызвать понос. Кроме того, Ж. к. обладают бактериостатическим действием.
Общая концентрация Ж. к. в крови и их соотношение существенно изменяются при ряде заболеваний печени и желчного пузыря, что используется в диагностических целях. При паренхиматозных поражениях печени резко снижается способность печеночных клеток захватывать Ж. к. из крови, в результате чего они накапливаются в крови и выделяются с мочой. Повышение концентрации Ж. к. в крови наблюдается и при затруднении оттока желчи, особенно при обтурации общего желчного протока (камень, опухоль), что сопровождается также нарушением печеночно-кишечной циркуляции с резким уменьшением или исчезновением конъюгатов дезоксихолатов из желчи. Длительное и существенное повышение концентрации Ж. к. в крови может оказывать повреждающее действие на печеночные клетки с развитием некрозов и изменением активности некоторых ферментов в сыворотке крови.
Высокая концентрация холатов в крови вызывает брадикардию и гипотонию, кожный зуд, гемолиз, повышение осмотической резистентности эритроцитов, нарушает процессы свертывания крови, замедляет скорость оседания эритроцитов. С выделением при болезнях печени Ж. к. через почки связывают развитие почечной недостаточности.
При острых и хрон, холециститах наблюдается уменьшение концентрации или полное исчезновение холатов из пузырной желчи, что объясняется уменьшением их образования в печени и ускорением всасывания их слизистой оболочкой воспаленного желчного пузыря.
Ж. к. и их производные разрушают в течение нескольких минут клетки крови, в том числе и лейкоциты, что следует учитывать при оценке диагностического значения количества лейкоцитов в дуоденальном содержимом. Холаты разрушают также и ткани, не соприкасающиеся в физиол, условиях с желчью, вызывают повышение проницаемости мембран и местное воспаление. При попадании желчи, напр., в брюшную полость быстро развивается тяжелый перитонит. В механизме развития острого панкреатита, антрального гастрита и даже язвы желудка определенную роль отводят Ж. к. Допускается возможность повреждения самого желчного пузыря конц. желчью, содержащей большое количество Ж. к. («химический» холецистит) .
Ж. к. являются исходным продуктом для производства стероидных гормонов. Благодаря сходству химического строения стероидных гормонов и Ж. к. последние обладают выраженным противовоспалительным действием. На этом свойстве Ж. к. основан метод лечения артритов местным применением конц. желчи (см. Желчь).
Для лечения поносов, возникающих после оперативного удаления части кишечника, и упорного кожного зуда у больных с заболеваниями печени и желчных путей применяются препараты, связывающие Ж. к. в кишечнике, напр, холестирамин.
См. также Жировой обмен, Печень.
Библиография: Комаров Ф. И. и Иванов А. И. Желчные кислоты, физиологическая роль, клиническое значение, Тер. арх., т. 44, № 3, с. 10, 1972; Куваева И. Б. Обмен веществ и кишечная микрофлора, М., 1976, библиогр.; Саратиков А. С. Желчеобразование и желчегонные средства, Томск, 1962; Успехи гепатологии, под ред. E. М. Тареева и А. Ф. Блюгера, в. 4, с. 141, Рига, 1973, библиогр.; Bergstrom S. a. Danielsson H. Formation and metabolisme of bile acids, Handb. Physiol., sect. 6, ed. by G. F. Code, p. 2391, Washington, 1968; The bile acids, chemistry, physiology and metabolism, ed. by P. P. Nair a. D. Kri-tshevsky, v. 1—2, N. Y., 1973, bibliogr.; Borgstrom B. Bile salts, Acta med. scand., v. 196, p. 1, 1974, bibliogr.; D a-nielsson H. a. S j o v a 1 1 J. Bile acid metabolism, Ann. Rev. Biochem., v. 44, p. 233, 1975, bibliogr.; Hanson R. F. a. o. Formation of bile acids in man, Biochim, biophys. Acta (Amst.), v. 431, p. 335, 1976; S h 1 у g i n G. K. Physiology of intestinal digestion, Progr, food Nutr., y. 2, p. 249, 1977, bibliogr.
Г. К. Шлыгин; Ф. И. Комаров (клин.).
Источник