Холестерин как предшественник в синтезе других стероидов
Содержание статьи
Обмен стероидов. Холестерин как предшественник ряда других стероидов. представление о биосинтезе холестерина — КиберПедия
Стероиды — производные восстановленных конденсированных циклических систем — циклопентанпергидрофенантренов. Холестерол — стероид, характерный только для животных организмов.
Холестерин усваивается из пищи—его эфиры гидролизуются холестеролэстеразой панкреатического или кишечного сока—продукты гидролиза всасываются в киш-ке в виде смешанных мицелл—экзогенный холестерин превращ в эфиры. В2 стадии:активация жир к-ты под действием ацетил-Коа-синтетазы; перенос ацильного остатка ацил-Коа на ОН-группу холестерина,катализирующий ацилхолестеролацилтрынсфераза—свободный холестерин упаковывается в хиломикроны,кот поступают в лимфу,потом в кровь—хиломикроны+ЛВП и получают белки(апоЕ-II)—в обратном направлении из хиломикрона в ЛВП поступают апоА-I и апоА-II.——хиломикроны превращ в в зрелые частицы—ТАГ расщепляются под действием липопротеинлипазы,прикрепляются к стенкам капилляров—обр остаточные хиломикроны——они удаляются из кров русла с помощью ЛНП-рецепторов,и расщепляются лизосомальными ферментами.—-Холестерин,освобожд.-ся из остаточных хиломикронов и липопротеинов,включаются в общий фонд этого стероида
В пр-се синтеза холестерина можно выделить 3 этапа:
1.Образование мевалоната из 3 остатков а-Коа.
а-Коа идет в цитоплазмув виде цитрата. В цитозоле кажд молекула ГМГ-Коа восстанавливается ГМГ-Коа-редуктазой в мевалонат с исп-м 2 NADPH+H
2. Образование сквалена 6 молекулами мевалоната.
мевалонат—изопентенилпирофосфаты—(образуется 30углеродное соединение сквалена)
3.Сквален превращается в холестерин.
Сквален циклизируется с обр полициклического ядра ланостерина—холестерин.
Выведение желчных кислот и холестерина из организма
Из организма человека ежедневно выводится около 1 г холестерола. Приблизительно половина этого количества экскретируется с фекалиями после превращения в желчные кислоты . Оставшаяся часть выводится в виде нейтральных стероид ов. Большая часть холестерола, поступившего в желчь, реабсорбируется; считается, что по крайней мере часть холестерола, являющегося предшественником фекальных стеролов , поступает из слизистой оболочки кишечника . Основным фекальным стеролом является копростанол , который образуется из холестерола в нижнем отделе кишечник а под действием присутствующей в нем микрофлоры . Значительная доля солей желчных кислот , поступающих с желчью, всасывается в кишечнике и через воротную вену возвращается в печень , где снова поступает в желчь. Этот путь транспорта солей желчных кислот получил название кишечно-печеночной циркуляции . Оставшаяся часть солей желчных кислот, а также их производные выводятся с фекалиями. Под действием кишечных бактерий первичные желчные кислоты превращаются во вторичные.
Регуляция обмена веществ. Гормоны.
Роль гормонов в системе регуляции метаболизма. Клетки-мишени и клеточные рецепторы гормонов.
Гормональные сигналы интегрируют и координируют метаболическую активность различных тканей, разносят информацию о размещении и распределении энергоемких веществ и веществ-предшественников синтеза биологических макромолекул к каждому органу.
Мишенью для гормона могут служить клетки одной или нескольких тканей. Воздействуя на клетку-мишень, гормон вызывает специфическую ответную реакцию. Характерный признак клетки-мишени — способность воспринимать информацию, закодированную в химической структуре гормона. Клетки-мишени отличают соответствующий гормон от множества других молекул и гормонов благодаря наличию на клетке-мишени соответствующего рецептора со специфическим центром связывания с гормоном. Рецепторы пептидных гормонов и адреналина располагаются на поверхности клеточной мембраны. Рецепторы стероидных и тиреоидных гормонов находятся внутри клетки. Причём внутриклеточные рецепторы для одних гормонов, например глюкокортикоидов, локализованы в цитозоле, для других, таких как андрогены, эстрогены, тиреоидные гормоны, расположены в ядре клетки. Рецепторы по своей химической природе являются белками и, как правило, состоят из нескольких доменов.
Источник
70.Холестерин как предшественник ряда других стероидов. Представление о биосинтезе холестерина. Написать ход реакций до образования мевалоновой кислоты. Роль гидроксиметилглутарил-КоА-редуктазы.
Холестерол
— стероид, характерный только для животных
организмов. Он синтезируется во многих
тканях человека, но основное место
синтеза — печень. В печени синтезируется
более 50% холестерола, в тонком кишечнике
— 15- 20%, остальной холестерол синтезируется
в коже, коре надпочечников, половых
железах. В сутки в организме синтезируется
около 1 г холестерола; с пищей поступает
300-500 мг Холестерол выполняет много
функций: входит в состав всех мембран
клеток и влияет на их свойства, служит
исходным субстратом в синтезе жёлчных
кислот и стероидных гормонов.
Предшественники в метаболическом пути
синтеза холестерола превращаются также
в убихинон — компонент дыхательной цепи
и долихол, участвующий в синтезе
гликопротеинов. Холестерол за счёт
своей гидроксильной группы может
образовывать эфиры с жирными кислотами.
Этерифицированный холестерол преобладает
в крови и запасается в небольших
количествах в некоторых типах клеток,
использующих его как субстрат для
синтеза других веществ. Холестерол и
его эфиры — гидрофобные молекулы, поэтому
они транспортируются кровью только в
составе разных типов ЛП. Обмен холестерола
чрезвычайно сложен — только для его
синтеза необходимо осуществление около
100 последовательных реакций. Всего в
обмене холестерола участвует около 300
разных белков. Нарушения обмена
холестерола приводят к одному из наиболее
распространённых заболеваний —
атеросклерозу. Смертность от последствий
атеросклероза (инфаркт миокарда, инсульт)
лидирует в общей структуре смертности
населения. Атеросклероз — «полигенное
заболевание», т.е. в его развитии
участвуют многие факторы, важнейшие из
которых наследственные. Накопление
холестерола в организме приводит к
развитию и другого распространённого
заболевания — желчнокаменной болезни.
Синтез
холестерола и егорегуляцияРеакции
синтеза холестерола происходят в
цитозоле клеток. Это один из самых
длинных метаболических путей в организме
человека.
Образованиемевалоната
Сложный
путь синтеза холестерола можно разделить
на 3 этапа
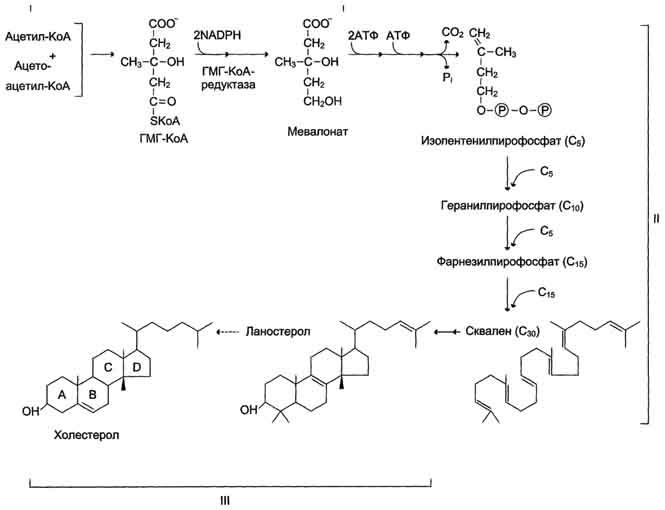
Первый
этап заканчивается образованием
мевалоната (мевалоновой кислоты). Две
молекулы ацетил-КоА конденсируются
ферментом тиолазой с образованием
ацетоацетил-КоА.
Фермент
щдроксиметилглутарил-КоА-синтаза
присоединяет третий ацетильный остаток
с образованием ГМГ-КоА
(3-гидрокси-3-метилглутарил-КоА). Эта
последовательность реакций сходна с
начальными стадиями синтеза кетоновых
тел:
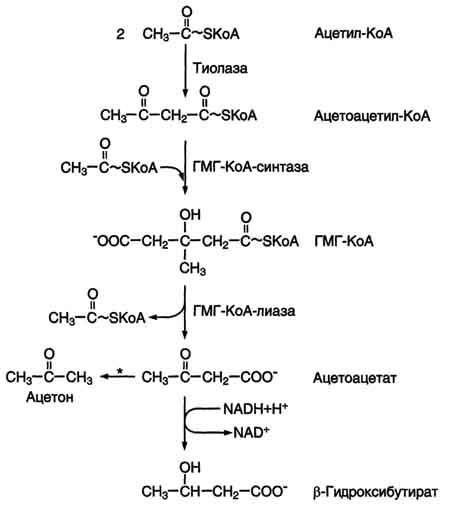
Следующая
реакция, катализируемая ГМГ-КоА-редуктазой,
является регуляторной в метаболическом
пути синтеза холестерола. В этой реакции
происходит восстановление ГМГ-КоА до
мевалоната с использованием 2 молекул
NADPH. Фермент ГМГ-КоА-редуктаза —
гликопротеин, пронизывающий мембрану
ЭР, активный центр которого выступает
в цитозоль.
Образование
скваленаНа
втором этапе синтеза мевалонат
превращается в пятиуглеродную
изопреноидную структуру, содержащую
пирофосфат — изопентенилпирофосфат.
Продукт конденсации 2 изопреновых единиц
— геранилпирофосфат. Присоединение ещё
1 изопреновой единицы приводит к
образованию фарнезилпирофосфата —
соединения, состоящего из 15 углеродных
атомов. Две молекулы фарнезилпирофосфата
конденсируются с образованием сквалена
— углеводорода линейной структуры,
состоящего из 30 углеродных атомов.
Образование
холестерола.На
третьем этапе синтеза холестерола
сквален через стадию образования
эпоксида ферментом циклазой превращается
в молекулу ланостерола, содержащую 4
конденсированных цикла и 30 атомов
углерода. Далее происходит 20 последовательных
реакций, превращающих ланостерол в
холестерол. На последних этапах синтеза
от ланостерола отделяется 3 атома
углерода, поэтому холестерол содержит
27 углеродных атомов.У холестерола
имеется насыщенная разветвлённая
боковая цепь из 8 углеродных атомов в
положении 17, двойная связь в кольце В
между атомами углерода в положениях 5
и 6, а также гидроксильная группа в
положении 3.В организме человека
изопентенилпирофосфат также служит
предшественником убихинона (KoQ) и
долихола, участвующего в синтезе
гликопротеинов.
Этерификация
холестерола.
В
некоторых тканях гидроксильная группа
холестерола этерифицируется с образованием
более гидрофобных молекул — эфиров
холестерола. Реакция катализируется
внутриклеточным ферментом АХАТ
(ацилКоА:холестеролаиилтрансферазой).
Реакция этерификации происходит также
в крови в ЛПВП, где находится фермент
ЛХАТ (лецитин:холестеролацилтрансфераза).
Эфиры холестерола — форма, в которой они
депонируются в клетках или транспортируются
кровью. В крови около 75% холестерола
находится в виде эфиров.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
21.05.2015184.29 Кб24Bazovaya_SLR_u_detey_33.pdf
- #
- #
- #
- #
- #
- #
- #
Источник
70.Холестерин как предшественник ряда других стероидов. Представление о биосинтезе холестерина. Написать ход реакций до образования мевалоновой кислоты. Роль гидроксиметилглутарил-КоА-редуктазы.
Холестерол
— стероид, характерный только для животных
организмов. Он синтезируется во многих
тканях человека, но основное место
синтеза — печень. В печени синтезируется
более 50% холестерола, в тонком кишечнике
— 15- 20%, остальной холестерол синтезируется
в коже, коре надпочечников, половых
железах. В сутки в организме синтезируется
около 1 г холестерола; с пищей поступает
300-500 мг Холестерол выполняет много
функций: входит в состав всех мембран
клеток и влияет на их свойства, служит
исходным субстратом в синтезе жёлчных
кислот и стероидных гормонов.
Предшественники в метаболическом пути
синтеза холестерола превращаются также
в убихинон — компонент дыхательной цепи
и долихол, участвующий в синтезе
гликопротеинов. Холестерол за счёт
своей гидроксильной группы может
образовывать эфиры с жирными кислотами.
Этерифицированный холестерол преобладает
в крови и запасается в небольших
количествах в некоторых типах клеток,
использующих его как субстрат для
синтеза других веществ. Холестерол и
его эфиры — гидрофобные молекулы, поэтому
они транспортируются кровью только в
составе разных типов ЛП. Обмен холестерола
чрезвычайно сложен — только для его
синтеза необходимо осуществление около
100 последовательных реакций. Всего в
обмене холестерола участвует около 300
разных белков. Нарушения обмена
холестерола приводят к одному из наиболее
распространённых заболеваний —
атеросклерозу. Смертность от последствий
атеросклероза (инфаркт миокарда, инсульт)
лидирует в общей структуре смертности
населения. Атеросклероз — «полигенное
заболевание», т.е. в его развитии
участвуют многие факторы, важнейшие из
которых наследственные. Накопление
холестерола в организме приводит к
развитию и другого распространённого
заболевания — желчнокаменной болезни.
Синтез
холестерола и егорегуляцияРеакции
синтеза холестерола происходят в
цитозоле клеток. Это один из самых
длинных метаболических путей в организме
человека.
Образованиемевалоната
Сложный
путь синтеза холестерола можно разделить
на 3 этапа
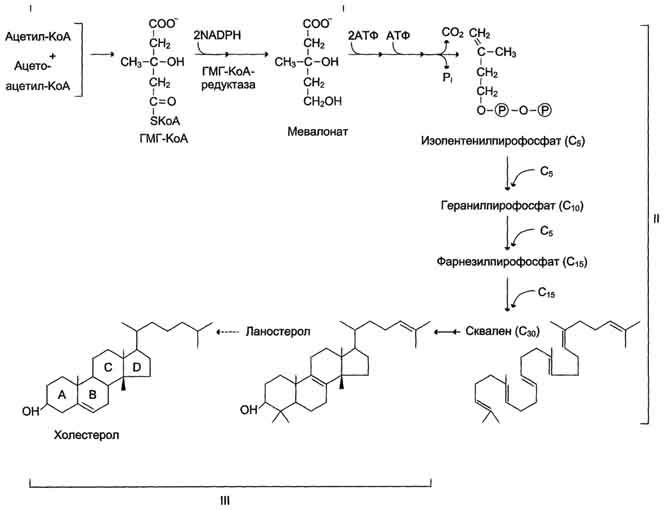
Первый
этап заканчивается образованием
мевалоната (мевалоновой кислоты). Две
молекулы ацетил-КоА конденсируются
ферментом тиолазой с образованием
ацетоацетил-КоА.
Фермент
щдроксиметилглутарил-КоА-синтаза
присоединяет третий ацетильный остаток
с образованием ГМГ-КоА
(3-гидрокси-3-метилглутарил-КоА). Эта
последовательность реакций сходна с
начальными стадиями синтеза кетоновых
тел:
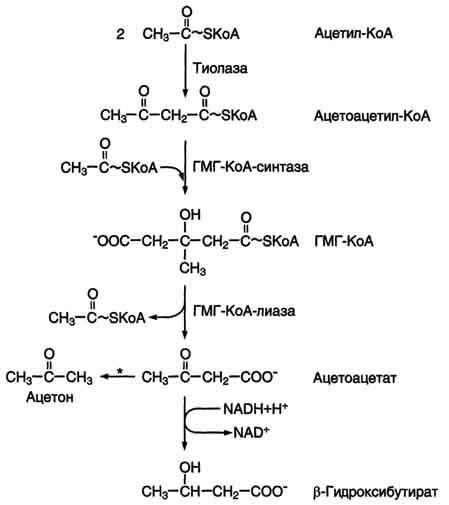
Следующая
реакция, катализируемая ГМГ-КоА-редуктазой,
является регуляторной в метаболическом
пути синтеза холестерола. В этой реакции
происходит восстановление ГМГ-КоА до
мевалоната с использованием 2 молекул
NADPH. Фермент ГМГ-КоА-редуктаза —
гликопротеин, пронизывающий мембрану
ЭР, активный центр которого выступает
в цитозоль.
Образование
скваленаНа
втором этапе синтеза мевалонат
превращается в пятиуглеродную
изопреноидную структуру, содержащую
пирофосфат — изопентенилпирофосфат.
Продукт конденсации 2 изопреновых единиц
— геранилпирофосфат. Присоединение ещё
1 изопреновой единицы приводит к
образованию фарнезилпирофосфата —
соединения, состоящего из 15 углеродных
атомов. Две молекулы фарнезилпирофосфата
конденсируются с образованием сквалена
— углеводорода линейной структуры,
состоящего из 30 углеродных атомов.
Образование
холестерола.На
третьем этапе синтеза холестерола
сквален через стадию образования
эпоксида ферментом циклазой превращается
в молекулу ланостерола, содержащую 4
конденсированных цикла и 30 атомов
углерода. Далее происходит 20 последовательных
реакций, превращающих ланостерол в
холестерол. На последних этапах синтеза
от ланостерола отделяется 3 атома
углерода, поэтому холестерол содержит
27 углеродных атомов.У холестерола
имеется насыщенная разветвлённая
боковая цепь из 8 углеродных атомов в
положении 17, двойная связь в кольце В
между атомами углерода в положениях 5
и 6, а также гидроксильная группа в
положении 3.В организме человека
изопентенилпирофосфат также служит
предшественником убихинона (KoQ) и
долихола, участвующего в синтезе
гликопротеинов.
Этерификация
холестерола.
В
некоторых тканях гидроксильная группа
холестерола этерифицируется с образованием
более гидрофобных молекул — эфиров
холестерола. Реакция катализируется
внутриклеточным ферментом АХАТ
(ацилКоА:холестеролаиилтрансферазой).
Реакция этерификации происходит также
в крови в ЛПВП, где находится фермент
ЛХАТ (лецитин:холестеролацилтрансфераза).
Эфиры холестерола — форма, в которой они
депонируются в клетках или транспортируются
кровью. В крови около 75% холестерола
находится в виде эфиров.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник
Обмен стероидов
Холестерол — основной стероид организма животных. У взрослого человека содержание холестерола составляет 140–150 г. Около 93% стероида входит в состав мембран и 7% находится в жидкостях организма. Холестерол увеличивает микровязкость мембран и снижает их проницаемость для Н 2 О и водорастворимых веществ. В крови он представлен в виде свободного холестерола, входящего в оболочку липопротеинов, и его эфиров, которые вместе с ТАГ составляют внутреннее содержимое этих частиц.
Холестериновое равновесие поддерживается благодаря тому, что с одной стороны холестерол поступает с пищей (~ 0,3 — 0,5 г ⁄ с ) и синтезируется в печени или других тканях (~ 0,5 г ⁄с ), а с другой — выводится с калом в виде желчных кислот, холестерола желчи, продуктов катаболизма стероидных гормонов, с кожным салом, в составе мембран слущенного эпителия (~ 1,0 г ⁄с ).
Холестерол является предшественником всех стероидов животного организма:
• желчных кислот, содержание которых у взрослого человека составляет около 5 г;
• стероидных гормонов: кортикостероидов, образующихся в корковом слое надпочечников, андрогенов — в семенниках и эстрагенов — в яичниках, синтез общего количества которых не превышает 40 мг ⁄с (с — сутки);
• витамина Д 3 , синтезирующегося в коже под действием УФО в количестве 10 мг ⁄с.
Ферменты синтеза холестерола имеются практически во всех клетках организма, но поскольку его концентрация в крови достаточно высока, а он способен репрессировать свой собственный синтез, то с заметной скоростью образование холестерола идет лишь в печени (~ 80%), слизистой кишечника (~ 10%), коре надпочечников, яичниках, семенниках и коже (~ 10%). В абсорбтивный период субстрат синтеза ацетил-КоА поступает из митохондрий в форме цитрата, когда в тканях образуется много АТФ, NADPH и ацетил-КоА в результате окисления глюкозы и жирных кислот. Первые стадии синтеза идут в цитозоле клеток, а более поздние на мембранах ЭПР .На первых стадиях процесса 18 молекул ацетил-КоА участвуют в синтезе 6 молекул ГМГ-КоА, в реакциях, сходных с реакциями синтеза кетоновых тел. Далее фермент ЭПРГМГ-КоА-редуктазавосстанавливает ГМГ-КоА вмевалоновую кислоту с затратой 2 молекул NADPH на каждый остаток ГМГ-КоА. Молекулы мевалоната фосфорилируются с помощью АТФ, декарбоксилируются и образуют 6 пятиуглеродных производных изопрена — изопентенилпирофосфаты. При последовательной конденсации этих соединений возникает симметричная линейная 30-углеродная молекула сквалена, которая через ряд стадий приобретает тетрациклическую структуруланостерола. Дальнейшие превращения ланостерола сопровождаются потерей трех метильных групп в виде СО 2 и образованием холестерола, содержащего 27 углеродных атомов, из которых 8 образуют углеводородную боковую цепь, два входят в метильные группы, а остальные 17 образуют циклопентанпергидрофенантреновую структуру.
Вопрос 35.Регуляция биосинтеза холестерина, транспорт холестерина кровью.
Ключевой регуляторный фермент — ГМГ-КоА-редуктаза, активность которого в печени регулируется трояким способом :
• на уровне транскрипции гена ГМГ-КоА-редуктазы. Корепрессорами процесса, снижающими скорость синтеза фермента, являются холестерол, желчные кислоты и кортикостероидные гормоны, а индукторами — инсулин и тиреоидные гормоны — Т3 и Т 4 ;
• путем фосфорилирования и дефосфорилирования, которое также регулируется гормонами. Дефосфорилирование стимулирует инсулин, который за счет активации протеинфосфатазы переводит фермент в дефосфорилированную активную форму, а глюкагон через аденилатциклазную систему обеспечивает механизм его фосфорилирования и инактивации;
• уменьшением количества фермента за счет протеолиза молекул, который стимулируют холестерол и желчные кислоты. Часть вновь синтезированного холестерола этерифицируется с образованием эфиров. Эту реакцию , как и в энтероцитах, катализирует АХАТ, присоединяя к холестеролу остатки линолевой или олеиновой кислот.
В транспорте холестерола и его эфиров по крови участвуют все липопротеины. Так, хиломикроны переносят холестерол из кишечника через кровь в печень в составе ХМост. В печени холестерол вместе с эндогенными жирами и фосфолипидами упаковывается в ЛПОНП и секретируется в кровь. В кровотоке ЛПОНП незрелые получают от ЛПВП мембранные белки АпоС II и АпоЕ и становятся зрелыми, т.е. способными взаимодействовать с ЛП-липазой, которая гидролизует ТАГ в составе ЛПОНП до ВЖК и глицерола. Частицы, теряя жиры, уменьшаются в размере, но возрастают по плотности и превращаются сначала в ЛППП, а затем в ЛПНП.
Date: 2015-05-22; view: 1249; Нарушение авторских прав
Источник