Как рассчитать кислородную емкость крови по гемоглобину
Содержание статьи
азы крови. Кислородная емкость крови и оксигемометрия
Газы крови. Кислородная емкость крови и оксигемометрия
Азот в крови находится только в растворенном виде, содержание его составляет в среднем 1,2 об. %. Будучи физиологически инертным, азот не участвует в обменных процессах, и измерение его содержания для изучения внешнего дыхания практического значения не имеет.
Так как основной задачей системы внешнего дыхания является поддержание на уровне констант содержания кислорода и углекислоты в крови, определение показателей, характеризующих газы крови, представляет значительный интерес для оценки функционального состояния легких.
В практической работе представляется полезным определение содержания и парциального давления кислорода и углекислоты в артериальной крови, поскольку содержание этих газов в венозной крови различных органов и даже в смешанной венозной крови, получаемой при катетеризации легочной артерии, зависит не столько от функции внешнего дыхания, сколько от уровня обмена веществ и скорости кровотока (Comroe с соавт.). Кислород, особенно углекислота, являются важнейшими компонентами буферных систем крови, вследствие чего исследование газов крови и кислотно-щелочного равновесия тесно взаимосвязаны.
Основными показателями, характеризующими газовый состав артериальной крови, считаются кислородная емкость крови в об.%, содержание кислорода и углекислоты в об.%, парциальное давление (напряжение) кислорода и углекислоты в миллиметрах ртутного столба, артерио-венозная разница по кислороду и венозно-артериальная разница по углекислоте в об.%. степень насыщения крови кислородом в процентах.
Кислородная емкость крови — максимальное количество кислорода, которое может быть связано кровью. В среднем 1 г гемоглобина связывает около 1,35 см3 кислорода. Поэтому кислородная емкость крови зависит не от функции внешнего дыхания, а от содержания гемоглобина. Содержание кислорода в крови также зависит не только от эффективности вентиляции, диффузии и газообмена в легких, но и от содержания гемоглобина в крови.
Весьма чувствительными показателями считаются парциальное давление (напряжение) кислорода и углекислоты. Определение парциального давления С02 может быть проведено вместе с измерением рН крови по методу Аструпа. Что же касается измерения парциального давления кислорода крови, то методика его сложна и вследствие этого не получила распространения в клинической практике.
Наибольшее практическое значение вполне оправданно отводится определению степени насыщения крови кислородом, методика которого в настоящее время значительно усовершенствована, и это исследование получило широкое распространение в клинике торакальной хирургии.
Методы определения степени насыщения крови кислородом разделяются на газометрические (манометрические) и оксигемометрические (спектрофотометрические). К первым относятся методы Ван-Слайка и Баркрофта. Газометрический способ Ван-Слайка основывается на принципах И. М. Сеченова — извлечение газов из крови в вакууме — и Холдейна — вытеснение газов химическими реактивами.
Исследование газов крови на аппарате Ван-Слайка получило широкое распространение в клинической физиологии. К числу достоинств этого метода относится высокая точность результатов и возможность определения содержания кислорода и углекислоты. Однако длительность и трудоемкость исследования, необходимость относительно большого количества крови (1 мл), для чего требуется пункция артерии, ограничивают его применение в практической работе легочного хирурга, тем более, что определение наиболее важного показателя — степени насыщения крови кислородом — в настоящее время обычно производится с помощью оксигемометрии — метода, значительно более доступного и мало уступающего в точности.
Поэтому показания к применению определения газов крови методом Ван-Слайка в легочной хирургии значительно ограничены, хотя он еще не потерял значения в исследовательской работе.
— Вернуться в оглавление раздела «Пульмонология.»
Оглавление темы «Спирометрия и оценка альвеолярного воздуха»:
1. Пробы с дозированными физическими нагрузкам. Спирография с физической нагрузкой
2. Раздельная спирография. Бронхоспирометрия
3. Недостатки бронхоспирометрии. Оценка ЖЕЛ каждого легкого
4. Искажение данных бронхоспирометрии. Оценка функциональных возможностей легкого
5. Проба с поочередным выключением легких из вентиляции. Однолегочная спирометрия
6. Проба с дозированной физической нагрузкой. Нагрузка при спирометрии
7. Проба с бронходилататорами. Общая и раздельная спирография во время операции на легких
8. Определение газового состава альвеолярного воздуха. Газоанализатор Холдейна
9. Раздельное исследование газового состава альвеолярного воздуха. Альвеолярный воздух при патологии
10. Газы крови. Кислородная емкость крови и оксигемометрия
Источник
ранспорт газов кровью. Транспорт кислорода. Кислородная емкость гемоглобина.
Оглавление темы «Вентиляция легких. Перфузия легких кровью.»:
1. Вентиляция легких. Вентиляция кровью легких. Физиологическое мертвое пространство. Альвеолярная вентиляция.
2. Перфузия легких кровью. Влияние гравитации на вентиляцию легких. Влияние гравитации на перфузию легких кровью.
3. Коэффициент вентиляционно-перфузионных отношений в легких. Газообмен в легких.
4. Состав альвеолярного воздуха. Газовый состав альвеолярного воздуха.
5. Напряжение газов в крови капилляров легких. Скорость диффузии кислорода и углекислого газа в легких. Уравнение Фика.
6. Транспорт газов кровью. Транспорт кислорода. Кислородная емкость гемоглобина.
7. Сродство гемоглобина к кислороду. Изменение сродства гемоглобина к кислороду. Эффект Бора.
8. Углекислый газ. Транспорт углекислого газа.
9. Роль эритроцитов в транспорте углекислого газа. Эффект Холдена..
10. Регуляция дыхания. Регуляция вентиляции легких.
Транспорт газов кровью. Транспорт кислорода. Кислородная емкость гемоглобина.
Кровообращение выполняет одну из важнейших функций переноса кислорода от легких к тканям, а углекислого газа — от тканей к легким. Потребление кислорода клетками тканей может изменяться в значительных пределах, например при переходе от состояния покоя к физической нагрузке и наоборот. В связи с этим кровь должна обладать большими резервами, необходимыми для увеличения ее способности переносить кислород от легких к тканям, а углекислый газ в обратном направлении.
Транспорт кислорода.
При 37 С растворимость 02 в жидкости составляет 0,225 мл • л-1 • кПа-1 (0,03 мл/л/мм рт. ст.). В условиях нормального парциального давления кислорода в альвеолярном воздухе, т. е. 13,3 кПа или 100 мм рт.ст., 1 л плазмы крови может переносить только 3 мл 02, что недостаточно для жизнедеятельности организма в целом. В покое в организме человека за минуту потребляется примерно 250 мл кислорода. Чтобы тканям получить такое количество кислорода в физически растворенном состоянии, сердце должно перекачивать за минуту огромное количество крови. В эволюции живых существ проблема транспорта кислорода была более эффективно решена за счет обратимой химической реакции с гемоглобином эритроцитов. Кислород переносится кровью от легких к тканям организма молекулами гемоглобина, которые содержатся в эритроцитах.
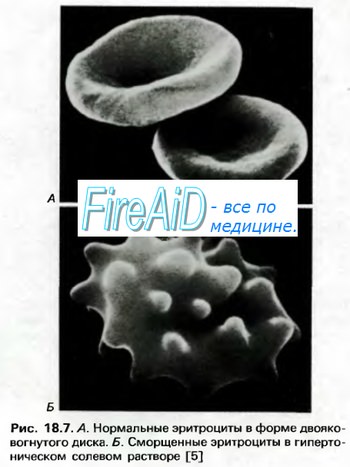
Гемоглобин способен захватывать кислород из альвеолярного воздуха (соединение называется ок-сигемоглобином) и освобождать необходимое количество кислорода в тканях. Особенностью химической реакции кислорода с гемоглобином является то, что количество связанного кислорода ограничено количеством молекул гемоглобина в эритроцитах крови. Молекула гемоглобина имеет 4 места связывания с кислородом, которые взаимодействуют таким образом, что зависимость между парциальным давлением кислорода и количеством переносимого кислорода с кровью имеет S-образную форму, которая носит название кривой насыщения или диссоциации оксигемоглобина (рис. 10.18). При парциальном давлении кислорода 10 мм рт. ст. насыщение гемоглобина кислородом составляет примерно 10 %, а при Р02 30 мм рт. ст. — 50—60 %. При дальнейшем увеличении парциального давления кислорода от 40 мм рт. ст. до 60 мм рт. ст. происходит уменьшение крутизны кривой диссоциации оксигемоглобина и процент его насыщения кислородом возрастает в диапазоне от 70—75 до 90 % соответственно. Затем кривая диссоциации оксигемоглобина начинает занимать практически горизонтальное положение, поскольку увеличение парциального давления кислорода с 60 до 80 мм рт. ст. вызывает прирост насыщения гемоглобина кислородом на 6 %. В диапазоне от 80 до 100 мм рт. ст. процент образования оксигемоглобина составляет порядка 2. В результате кривая диссоциации оксигемоглобина переходит в горизонтальную линию и процент насыщения гемоглобина кислородом достигает предела, т. е. 100. Насыщение гемоглобина кислородом под влиянием Р02 характеризует своеобразный молекулярный «аппетит» этого соединения к кислороду.
Значительная крутизна кривой насыщения гемоглобина кислородом в диапазоне парциального давления от 20 до 40 мм рт. ст. способствует тому, что в ткани организма значительное количество кислорода может диффундировать из крови в условиях фадиента его парциального давления между кровью и клетками тканей (не менее 20 мм рт. ст.). Незначительный процент насыщения гемоглобина кислородом в диапазоне его парциального давления от 80 до 100 мм рт. ст. способствует тому, что человек без риска снижения насыщения артериальной крови кислородом может перемещаться в диапазоне высот над уровнем моря до 2000 м.
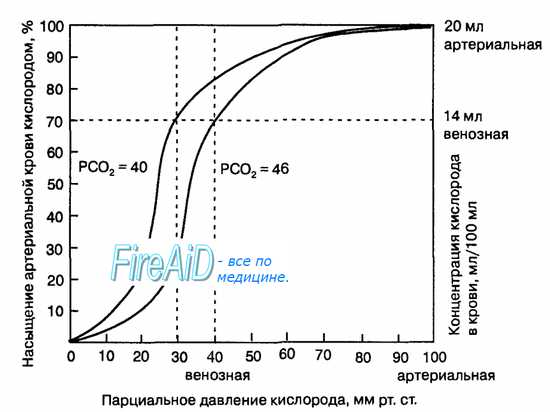
Рис. 10.18. Кривая диссоциации оксигемоглобина. Пределы колебания кривой при РС02 = 40 мм рт. ст. (артериальная кровь) и РС02 = 46 мм рт. ст. (венозная кровь) показывают изменение сродства гемоглобина к кислороду (эффект Ходена).
Общие запасы кислорода в организме обусловлены его количеством, находящимся в связанном состоянии с ионами Fe2+ в составе органических молекул гемоглобина эритроцитов и миоглобина мышечных клеток.
Один грамм гемоглобина связывает 1,34 мл 02. Поэтому в норме при концентрации гемоглобина 150 г/л каждые 100 мл крови могут переносить 20,0 мл 02.
Количество 02, которое может связаться с гемоглобином эритроцитов крови при насыщении 100 % его количества, называется кислородной емкостью гемоглобина. Другим показателем дыхательной функции крови является содержание 02 в крови (кислородная емкость крови), которое отражает его истинное количество, как связанного с гемоглобином, так и физически растворенного в плазме. Поскольку в норме артериальная кровь насыщена кислородом на 97 %, то в 100 мл артериальной крови содержится примерно 19,4 мл 02.
— Также рекомендуем «Сродство гемоглобина к кислороду. Изменение сродства гемоглобина к кислороду. Эффект Бора.»
Источник
Количество газов в крови (мл/л)
| кровь | О2 | СО2 |
| артериальная | 180-200 | |
| венозная | 130-150 |
Кислородная емкость крови (КЕК) – максимальное количество кислорода, которое может быть связано 1 л крови. Составляет 180-200 мл/л (табл. 4).
Кислородную емкость крови определяют по концентрации Нb с двухвалентным железом (Fe2+). Максимально 1 моль гемоглобина может присоединить 4 моля О2 к своим 4 молям железа гема. Принимая во внимание молекулярную массу (64500 Да) 1 г гемоглобина связывает 4/64500 = 0,062 ммоль О2. Так как молярный объем идеального газа равен 22,4 л/моль, то 1г гемоглобина связывает 1,39 мл О2 (0,062´22,4).
Измерения непосредственно в крови демонстрируют меньшую величину, так как некоторая часть гемоглобина в организме в нормальных условиях находится в измененной форме, которая не может связываться с О2 (например карбоксигемоглобин НbCО, метгемоглобин). Для практических целей применяют величину, называемую числом Гюфнера, приблизительно равную 1,34 мл О2 на 1 г Нb.
Коэффициент утилизации кислорода (КУО2)представляет собой часть кислорода, поглощаемую тканями из капиллярного русла.
КУО2 —это процентное отношение доли кислорода, используемой тканями (разности концентраций кислорода в артериальной и венозной крови), к общему содержанию его в артериальной крови:
КУО2 = VO2а- VO2в / VO2а x 100 %
Скорость доставки кислорода в нормальных условиях значительно превышает его потребление, в результате чего лишь малая доля доступного кислорода извлекается из капиллярной крови в обычном состоянии (в покое КУО2 = 25-35%). Это позволяет тканям приспосабливаться к снижению доставки кислорода
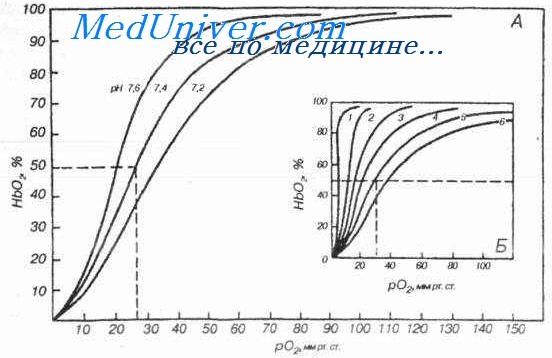
Зависимость степени оксигенации гемоглобина от парциального давления О2 в альвеолярном воздухе графически представляется в виде кривой диссоциации оксигемоглобина, или сатурационной кривой (рис. 15).
Рис. 15. Кривая диссоциации оксигемоглобина, или сатурационная кривая
Плато кривой диссоциации характерно для насыщенной О2 (сатурированной) артериальной крови, а крутая нисходящая часть кривой — венозной, или десатурированной, крови в тканях. Кривая имеет S-образную форму, что обусловлено кооперативным взаимодействием четырех субъединиц, составляющих тетрамер гемоглобина. Присоединение О2 к гемму одной субъединицы повышает афинность (сродство) для его соединения с остальными субъединицами. Важным параметром для описания этой кривой является парциальное давление О2 при половинном насыщении гемоглобина. В человеческой крови Р0,5=27 мм рт.ст.
S-образная форма кривой связывания гемоглобином О2 имеет большое значение для транспортной функции крови. В области значений выше 65 мм рт.ст. кривая плоская, изменение Ро2 только немного меняет насыщение кислородом. Это область нормальных значений альвеолярного Ро2, которое может немного снижаться без заметного уменьшения насыщения гемоглобина кислородом в крови капилляров легких (например при подъеме в горы). Отвесный спад в нижней части кривой связывания (менее 65 мм рт.ст.) гарантирует, что Ро2 в капиллярной крови периферических тканей, несмотря на отдачу кислорода, остается достаточно высоким, чтобы обеспечить ткани кислородом путем диффузии.
Ряд факторов влияет на афинность (сродство) гемоглобина к О2. При этом в первую очередь изменяется положение кривой и гораздо меньше – ее форма. Различные метаболические факторы вызывают смещения кривой диссоциации влево или вправо (рис. 16).
ВЛЕВО ВПРАВО
Рис. 16. Сдвиги кривой диссоциации оксигемоглобина
Сродство гемоглобина к кислороду регулируется важнейшими факторами метаболизма тканей: Ро2 pH, температурой и внутриклеточной концентрацией 2,3-дифосфоглицерата.
1) Величина рН и содержание СО2 в любой части организма закономерно изменяют сродство гемоглобина к О2: уменьшение рН крови вызывает сдвиг кривой диссоциации соответственно вправо (уменьшается сродство гемоглобина к О2), а увеличение рН крови — сдвиг кривой диссоциации влево (повышается сродство гемоглобина к О2). Влияние рН на кривую диссоциации оксигемоглобина называется «эффектом Бора». Эффект Бора может быть связан также с изменением Рсо2: его повышение сдвигает кривую связывания О2 вправо, понижение — влево. Это влияние основывается, в первую очередь, на том, что повышение Рсо2 ведет к снижению рH и наоборот. Влияние самих молекул СО2 на афинность гемоглобина к О2, напротив, незначительно.
2) Влияние температуры: рост температуры уменьшает сродство гемоглобина к О2. В работающих мышцах увеличение температуры способствует освобождению О2. Уменьшение температуры тканей вызывает сдвиг влево кривой диссоциации оксигемоглобина. Значение влияния температуры для гомойотермных организмов небольшое, хотя температура крови на периферии тела может сильно отличаться от температуры всего организма.
3) Эффект 2,3-дифосфоглицерата:в его отсутствии афинность гемоглобина к кислороду очень высока. Главная функция 2,3-дифосфоглицерата состоит в смещении кривой связывания О2 вправо, в область, которая соответствует физиологическим условиям.
Контрольные вопросы
1. Какие транспортные формы кислорода существуют ?
2. Что такое кислородная емкость крови, как она определяется ?
3. Что такое коэффициент утилизации кислорода, как он определяется ?
4. Почему кривая диссоциации оксигемоглобина (сатурационная кривая) имеет S- образную форму ?
5. Какое биологическое значение крутой и отлогой частей кривой диссоциации оксигемоглобина (сатурационной кривой) ?
6. Какие факторы влияют на афинность (сродство) гемоглобина к О2 ?
Источник
емодинамические параметры. Кислородный поток и потребление кислорода
Гемодинамические параметры. Кислородный поток и потребление кислородаИз многочисленных гемодинамических параметров, ответственных за транспорт кислорода, мы остановимся только на тех, которые могут быть реализованы в мониторинге кислородного баланса организма с помощью стандартной аппаратуры. Приоритетной задачей мониторинга кислородного баланса организма является регистрация двух основных параметров: Оба эти параметра могут быть легко рассчитаны, если имеются сведения о минутном объеме сердца, концентрации гемоглобина и количестве оксигемоглобина в артериальной и венозной крови. Нb — концентрация гемоглобина (г/л),
В клинической физиологии принято отражать истинные величины параметров в виде индексов, выражающих величину параметра с учетом поверхности тела. Поэтому параметры кислородного баланса здорового человека массой 75 кг выражаются следующими величинами: Из этих простых расчетов можно сделать одно очень важное заключение. В состоянии покоя кислородный поток более чем в 3,5 раза превышает потребление кислорода, а отношение потребления 02 к его потоку V02 / D02 равняется 0,28. А это значит, что только 1/3 кислородного потока расходуется на энергетические затраты организма, а 70-75% кислорода возвращается в венозную систему (Pv02 составляет около 40 мм Нд). Следовательно, сатурация венозной крови или артерио-венозное различие по кислороду (a-v D02) является еще одним важным критерием транспорта кислорода, указывающим на соответствие транспортируемого объема кислорода его потребностям для обеспечения тканевого метаболизма. При гиперметаболических состояниях увеличение потока (транспорта кислорода) реализуется за счет повышения функции сердечно-сосудистой системы. Возрастает частота сердечных сокращений, достигая в некоторых случаях 180-190 циклов/мин. Ударный объем сердца может повыситься до 130-140 мл, сердечный выброс — до 25-30 л/мин. При этих состояниях подсоединяется еще один компенсаторный механизм — в кровоток включаются ранее нефункционирующие капилляры, которые в состоянии покоя составляют до 75-80%. И, наконец, как было указано выше, не последнюю роль в компенсации повышенного потребления кислорода играет и гемический фактор: смещение кривой диссоциации оксигемоглобина вправо, облегчающее поступление кислорода в ткани. В этом плане вызывают определенный интерес рекомендации некоторых авторов рассчитывать еще один интегральный параметр кривой диссоциации оксигемоглобина. Речь идет о показателе, называемом «давление извлечения кислорода — Рх», который выражается величиной напряжения кислорода, достаточной для извлечения 2,3 ммоль (51 мл) его из 1 литра крови. Этот интегральный показатель, устанавливая взаимовлияния Ра02, кислородной емкости крови и аффинитета гемоглобина к кислороду, отражает суммарный эффект этих факторов по влиянию на доставку кислорода к тканям. В норме он соответствует 38 мм Нg. Снижение Рх указывает на дефицит кислородного потока. Мы намеренно приводим этот показатель, т.к. с его помощью можно получить ориентировочное представление о кислородном потоке в случаях, когда не удается его рассчитать по формуле, например, при отсутствии возможности регистрировать сердечный выброс. — Также рекомендуем «Факторы влияющие на потребление кислорода. Предупреждение критической тканевой гипоксии» Оглавление темы «Параметры потребления кислорода и гемодинамики»: |
Источник