Кривая насыщения кислородом гемоглобин
Содержание статьи
Газы крови влияют на pH
Несмотря на то, что исследование кислотно-основного состояния, строго говоря, подразумевает исследование только величины pH (концентрации ионов H+), в реальности в него также включается исследование физиологически важных газов, присутствующих в крови – O2 и CO2. Анализ газов показывает эффективность газообмена по величинам парциальных давлений – pO2 и pCO2.
Через альвеолярную мембрану молекулы любых газов перемещаются диффузно по градиенту концентрации. Молекулы O2 атмосферного воздуха поступают из альвеол в кровь, а молекулы CO2 из крови в альвеолы до тех пор пока их парциальные давления не выровняются.
Величина парциального давления – это процентная доля газа в общем объеме.
Углекислый газ
Концентрация СО2 в альволярном воздухе столь низка, а в крови столь высока, что диффузия этого газа в альвеолы чрезвычайно эффективна и скорость его удаления зависит только от альвеолярной вентиляции – общего объема воздуха, транспортируемого в минуту между альвеолами и атмосферой («скорости выдувания»).
Следовательно,
- при усиленной вентиляции легких углекислый газ быстро выводится, и показатель pCO2в крови снижается. Это означает потерю организмом угольной кислоты (ионов H+), что является причиной защелачивания крови – алкалоза, называемого дыхательным или респираторным.
- при недостаточной альвеолярной вентиляции величина рСО2 повышается, что свидетельствует о недостаточном его удалении и накоплении H2CO3. Иными словами, повышение в крови показателя рСО2 является причиной дыхательного ацидоза.
Увеличенное pCO2 (гиперкапния) всегда свидетельствует о снижении альвеолярной вентиляции.
Кислород
Вопросы, связанные с оксигенацией крови и транспортом кислорода более сложны. Связано это с тем, что в виде свободных (растворенных) молекул O2 находится лишь небольшая доля общего кислорода крови. Основная часть кислорода связана с гемоглобином (оксигемоглобин) и истинное содержание кислорода зависит от двух дополнительных параметров – концентрации Hb и насыщения (сатурации) гемоглобина кислородом.
Оксигемоглобин
Оксигемоглобин (HbО2) – процентное содержание в крови, является отношением фракции оксигемоглобина (HbО2) к сумме всех фракций (общему гемоглобину).
Насыщение гемоглобина кислородом
Насыщение гемоглобина кислородом (HbOSAT, SО2), представляет собой отношение фракции оксигенированного гемоглобина к тому количеству гемоглобина в крови, который способен транспортировать О2.
Отличия между двумя показателями HbО2 и HbOSAT заключаются в том, что у пациентов возможно наличие в крови такой формы гемоглобина, которая не способна акцептировать О2 (Hb‑CO, metHb, сульфоHb). Но так как большинство больных не имеют в крови повышенного содержания этих форм гемоглобина, значения HbО2 и SО2 обычно очень близки.
Например, если при отравлении нитритами количество metHb составляет 15%, тогда величина HbО2 никогда не сможет превысить 85%, но насыщение (HbOsat) может быть различно – от максимума (HbOsat=95-98%) при полном насыщении до низких величин при отсутствии кислорода.
Показатель насыщения кислородом показывает процент доступных мест связывания на гемоглобине.
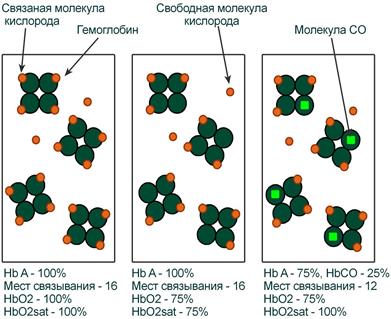
Иллюстрация понятий оксигемоглобина (HbO2) и насыщения гемоглобина (HbO2sat)
Парциальное давление кислорода (pO2)
Парциальное давление O2 выступает как движущая сила, приводящая к насыщению гемоглобина кислородом. И хотя, как правило, чем выше pO2 тем выше HbOsat, эта зависимость не является линейной.
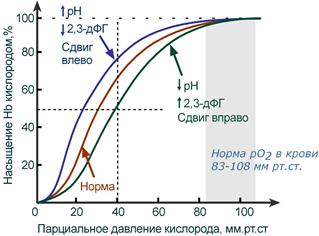
Кривая диссоциации гемоглобина в норме и
при изменении pH и концентрации 2,3-дифосфоглицерата
В центральной части кривой насыщения (или кривой диссоциации) гемоглобина малейшие сдвиги pO2 приводят к резким изменениям насыщения гемоглобина. И наоборот, при высоком pO2 (80-90-100 мм рт.ст) кривая становится плоской, насыщение гемоглобина мало зависит от колебаний кислорода в плазме.
Сдвиг влево происходит при защелачивании и снижении концентрации 2,3-дифосфоглицерата и сигнализирует об увеличении сродства кислорода к гемоглобину (в легких). Сдвиг вправо — это снижение сродства кислорода к гемоглобину (в тканях), обеспечивается закислением среды и накоплением 2,3-дифосфоглицерата.
Показатель pO2 не отражает содержание кислорода в цельной крови! Но хотя pO2 и не показывает общее количество кислорода в крови, но это общее количество зависит от pO2 через показатель сатурации гемоглобина.
В свою очередь имеются факторы, влияющие на величину pO2:
1. Альвеолярная вентиляция. Хотя она влияет как на pO2 так и на pCO2, но доля кислорода в альвеолах при гипервентиляции может лишь слегка увеличиться, приближаясь к pO2 атмосферного воздуха, при гиповентиляции – стремительно падает, вытесняясь поступающим из крови CO2. В то же время доля CO2 в альвеолах быстро снижается при усиленной вентиляции.
2. Вентиляционно-перфузионное соотношение, определяется тем, что
- не вся кровь, притекающая к легким, соприкасается с хорошо вентилируемыми альвеолами (спадение альвеол, уплотнение стенки).
- не все хорошо вентилируемые альвеолы получают достаточно крови (правожелудочковая сердечная недостаточность).
3. Концентрация кислорода во вдыхаемом воздухе (FiO2, fraction of inspired oxygen).
В таблице приведены сравнительные величины концентрации кислорода и углекислого газа в воздухе, крови и тканях.
Необходимо обратить внимание на перепады концентраций кислорода и углекислого газа в крови и альвеолярном воздухе. Важной особенностью является то, что pO2 в альвеолярном воздухе и артериальной крови очень близки, т.е. в обычных условиях глубоким и/или частым дыханием невозможно повысить потребление кислорода и насыщение им гемоглобина. В то же время разность концентраций pCO2 в венозной крови и альвеолярном воздухе позволяет эффективно его удалять при частом дыхании.
| pO2, мм рт.ст. | pCO2, мм рт.ст | |
| Вдыхаемый воздух | 159 | 0,23 |
| Альвеолярный воздух | 105-110 | 40 |
| Артериальная кровь | 83-108 | 35-45 |
| Ткани | 10-20 | 50-60 |
| Венозная кровь | 35-49 | 46-51 |
| Выдыхаемый воздух | 116 | 32 |
Источник
родство гемоглобина к кислороду. Изменение сродства гемоглобина к кислороду. Эффект Бора.
Оглавление темы «Вентиляция легких. Перфузия легких кровью.»: Сродство гемоглобина к кислороду. Изменение сродства гемоглобина к кислороду. Эффект Бора.Молекула гемоглобина может находиться в двух формах — напряженной и расслабленной. Расслабленная форма гемоглобина имеет свойство насыщаться кислородом в 70 раз быстрее, чем напряженная. Изменение фракций напряженной и расслабленной формы в общем количестве гемоглобина в крови обусловливает S-образный вид кривой диссоциации оксигемоглобина, а следовательно, так называемое сродство гемоглобина к кислороду. Если вероятность перехода от напряженной формы гемоглобина к расслабленной больше, то возрастает сродство гемоглобина к кислороду, и наоборот. Вероятность образования указанных фракций гемоглобина изменяется в большую или меньшую сторону под влиянием нескольких факторов. Основной фактор — это связывание кислорода с геминовой фуппой молекулы гемоглобина. При этом чем больше геминовых фупп гемоглобина связывают кислород в эритроцитах, тем более легким становится переход молекулы гемоглобина к расслабленной форме и тем выше их сродство к кислороду. Поэтому при низком Р02, что имеет место в метаболически активных тканях, сродство гемоглобина к кислороду ниже, а при высоком Р02 — выше. Как только гемоглобин захватывает кислород, повышается его сродство к кислороду и молекула гемоглобина становится насыщенной при связывании с четырьмя молекулами кислорода.
Когда эритроциты, содержащие гемоглобин, достигают тканей, то кислород из эритроцитов диффундирует в клетки. В мышцах он поступает в своеобразного депо кислорода — в молекулы миоглобина, из которого кислород используется в биологическом окислении мышц. Диффузия кислорода из гемоглобина эритроцитов в ткани обусловлена низким Р02 в тканях — 35 мм рт. ст. Внутри клеток тканей напряжение кислорода, необходимое для поддержания нормального метаболизма, составляет еще меньшую величину — не более 1 кПа. Поэтому кислород путем диффузии из капилляров достигает метаболически активных клеток. Некоторые ткани приспособлены к низкому содержанию Р02 в капиллярах крови, что компенсируется высокой плотностью капилляров на единицу объема тканей. Например, в скелетной и сердечной мышцах Р02 в капиллярах может снизиться чрезвычайно быстро во время сокращения. В мышечных клетках содержится белок миоглобин, который имеет более высокое сродство к кислороду, чем гемоглобин. Миоглобин интенсивно насыщается кислородом и способствует его диффузии из крови в скелетную и сердечную мышцы, где он обусловливает процессы биологического окисления. Эти ткани способны экстрагировать до 70 % кислорода из крови, проходящей через них, что обусловлено снижением сродства гемоглобина к кислороду под влиянием температуры тканей и рН. Эффект рН и температуры на сродство гемоглобина к кислороду. Молекулы гемоглобина способны реагировать с ионами водорода, в результате этой реакции происходит снижение сродства гемоглобина к кислороду. При насыщении гемоглобина менее 100 % низкое рН понижает связывание кислорода с гемоглобином — кривая диссоциации оксигемоглобина смещается вправо по оси х. Это изменение свойства гемоглобина под влиянием ионов водорода называется эффектом Бора. Метаболически активные ткани продуцируют кислоты, такую как молочная, и С02. Если рН плазмы крови снижается от 7,4 в норме до 7,2, что имеет место при сокращении мыщц, то концентрация кислорода в ней будет возрастать вследствие эффекта Бора. Например, при постоянном рН 7,4 кровь отдавала бы порядка 45 % кислорода, т. е. насыщение гемоглобина кислородом снижалось до 55 %. Однако когда рН снижается до 7,2, кривая диссоциации смещается по оси х вправо. В результате насыщение гемоглобина кислородом падает до 40 %, т. е. кровь может отдавать в тканях до 60 % кислорода, что на 1/з больше, чем при постоянном рН.
Метаболически активные ткани повышают продукцию тепла. Повышение температуры тканей при физической работе изменяет соотношение фракций гемоглобина в эритроцитах и вызывает смещение кривой диссоциации оксигемоглобина вправо вдоль оси х. В результате большее количество кислорода будет освобождаться из гемоглобина эритроцитов и поступать в ткани. Эффект 2,3-дифосфоглицерата (2,3-ДФГ) на сродство гемоглобина к кислороду. При некоторых физиологических состояниях, например при понижении Р02 в крови ниже нормы (гипоксия) в результате пребывания человека на большой высоте над уровнем моря, снабжение тканей кислородом становится недостаточным. При гипоксии может понижаться сродство гемоглобина к кислороду вследствие увеличения содержания в эритроцитах 2,3-ДФГ. В отличие от эффекта Бора, уменьшение сродства гемоглобина к кислороду под влиянием 2,3-ДФГ не является обратимым в капиллярах легких. Однако при движении крови через капилляры легких эффект 2,3-ДФГ на снижение образования оксигемоглобина в эритроцитах (плоская часть кривой диссоциации оксигемоглобина) выражен в меньшей степени, чем отдача кислорода под влиянием 2,3-ДФГ в тканях (наклонная часть кривой), что обусловливает нормальное кислородное снабжение тканей. — Также рекомендуем «Углекислый газ. Транспорт углекислого газа.» |


Источник
иссоциация оксигемоглобина и ее зависимость. Эффект Бора
Диссоциация оксигемоглобина и ее зависимость. Эффект Бора
а) Факторы, сдвигающие кривую диссоциации оксигемоглобина. Их значение для транспорта кислорода. Кривые диссоциации оксигемоглобина (для облегчения понимания просим вас изучить рисунки ниже) справедливы для нормальной крови со средними показателями. Однако существует ряд факторов, которые могут сдвигать эту кривую в одну или другую сторону. На рисунке видно, что при некотором закислении крови со снижением рН от нормального уровня 7,4 до 7,2 кривая диссоциации смещается в среднем на 15% вправо, а повышение уровня рН от нормального уровня 7,4 до 7,6 смещает кривую на такое же расстояние влево.
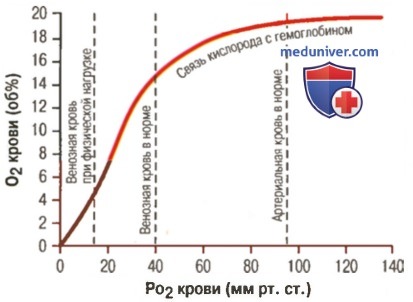
Влияние PO2 крови на количество связанного гемоглобина на 100 мл крови
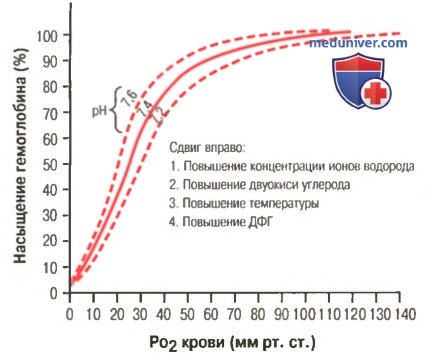
Сдвиг кривой диссоциации оксигемоглобина вправо в результате повышения концентрации ионов водорода (снижения pH). ДФГ — 2,3-дифосфоглицерат
Кроме изменений рН известны и другие факторы, которые могут сдвигать кривую диссоциации. Назовем три, действие которых сдвигает кривую вправо: (1) повышение концентрации двуокиси углерода; (2) повышение температуры крови; (3) повышение концентрации 2,3-дифосфоглицерата — метаболически важного фосфата, который в зависимости от метаболических условий присутствует в крови в разных концентрациях.
б) Повышение снабжения тканей кислородом в случаях, когда двуокись углерода и ионы водорода сдвигают кривую диссоциации оксигемоглобина. Эффект Бора. Сдвиг кривой диссоциации оксигемоглобина в ответ на повышение содержания двуокиси углерода и ионов водорода в крови имеет существенное влияние, выражающееся в ускорении высвобождения кислорода из крови в тканях и увеличении оксигенации крови в легких. Это называют эффектом Бора и объясняют его следующим образом.
При прохождении крови через ткань двуокись углерода диффундирует из клеток ткани в кровь. В результате в крови увеличивается PO2, а затем концентрации угольной кислоты (H2CO3) и ионов водорода. Эти изменения сдвигают кривую диссоциации оксигемоглобина вправо и вниз (для облегчения понимания просим вас изучить рисунок выше), уменьшая сродство кислорода к гемоглобину, и в результате увеличивается выход кислорода в ткани.
При диффузии двуокиси углерода из крови в альвеолы происходят процессы обратного направления — в результате в крови снижаются PCO2 и концентрация ионов водорода, сдвигая кривую диссоциации оксигемоглобина влево и вверх. При этом значительно увеличивается количество кислорода, связывающегося с гемоглобином при любом существующем уровне альвеолярного PO2, что увеличивает транспорт кислорода к тканям.
в) Сдвиг кривой диссоциации оксигемоглобина под влиянием дифосфоглицерата. Нормальное содержание ДФГ в крови вызывает постоянный небольшой сдвиг кривой диссоциации оксигемоглобина вправо. В случае гипоксического состояния, продолжающегося более нескольких часов, концентрация ДФГ в крови значительно возрастает, и кривая диссоциации оксигемоглобина сдвигается вправо еще больше.
В присутствии такой концентрации ДФГ кислород в тканях высвобождается при PO2, превышающем нормальный уровень на 10 мм рт. ст., поэтому в некоторых случаях такой механизм с участием ДФГ может оказаться важным для адаптации к гипоксии, особенно если причиной гипоксии является уменьшение в ткани кровотока.
г) Сдвиг кривой диссоциации во время физической нагрузки. Во время физической нагрузки некоторые факторы вызывают значительный сдвиг кривой диссоциации оксигемоглобина вправо, поэтому активные, выполняющие физическую работу мышечные волокна получают дополнительное количество кислорода. В свою очередь, работающие мышцы высвобождают большое количество двуокиси углерода; это в совокупности с действием некоторых других кислот, высвобождающихся мышцами, повышает концентрацию ионов водорода в крови капилляров мышц.
Кроме того, во время работы температура мышцы часто повышается на 2-3°С, что может еще больше увеличивать доставку кислорода мышечным волокнам. Все эти факторы вызывают значительный сдвиг кривой диссоциации оксигемоглобина в крови капилляров мышц вправо. Сдвиг вправо означает высвобождение кислорода гемоглобином в мышце при достаточно высоком уровне PO2 (40 мм рт. ст.) даже в случаях, когда из него уже высвободилось 70% кислорода. Сдвиг кривой в другую сторону показывает, что в легких присоединилось дополнительное количество кислорода из альвеолярного воздуха.
— Также рекомендуем «Участие кислорода в метаболизме. Метаболическое потребление кислорода»
Оглавление темы «Кислород и его доставка в организме»:
1. Диффузия газов через дыхательную мембрану. Дыхательная мембрана
2. Емкость дыхательной мембраны. Диффузионная емкость для кислорода
3. Вентиляционно-перфузионный коэффициент. Парциальное давление кислорода и двуокиси углерода
4. Концепция физиологического шунта. Концепция физиологического мертвого пространства
5. Обмен кислорода в организме. Транспорт кислорода из легких в ткани
6. Транспорт кислорода артериальной кровью. Диффузия кислорода
7. Гемоглобин. Роль гемоглобина в транспорте кислорода
8. Коэффициент использования кислорода. Сохранение постоянства кислорода в тканях
9. Диссоциация оксигемоглобина и ее зависимость. Эффект Бора
10. Участие кислорода в метаболизме. Метаболическое потребление кислорода
Источник
Физиология и патофизиология газообмена
Транспорт О2 из атмосферы до периферических тканей
организма представляет собой многоступенчатый процесс.
После того как воздух попадает в альвеолы легких, следующим этапом
газообмена является диффузия кислорода из альвеол в кровь легочных
капилляров и диффузия СО2 из крови легочных капилляров
в альвеолы. Диффузия представляет собой простое движение молекул через
респираторную мембрану из области более высокого давления в область
более низкого.
Таким образом, парциальное давление О2 (PO2)
является одним из основных факторов, определяющих его транспорт в
организме, причем не только в легких, но и во всем теле вплоть до
периферических тканей.
Помимо градиента давления скорость диффузии определяется 1) растворимостью
газа в жидкости; 2) площадью поверхности, через которую протекает
диффузия; 3) расстоянием, которое газ должен пройти при диффузии;
4) молекулярным весом газа; 5) температурой жидкости. Поскольку в
живом организме температура постоянна, она обычно не учитывается.
Применительно к альвеолокапиллярной мембране скорость диффузии газа
будет зависеть от:
- толщины мембраны;
- площади поверхности мембраны;
- диффузионного коэффициента газа в мембране;
- градиента давления газа по обе стороны мембраны.
По мере транспорта О2 от легких к периферическим
тканям его парциальное давление снижается. Если в атмосферном воздухе
при нормальном атмосферном давлении парциальное давление О2
составляет 159 мм рт. ст., то в периферических тканях, в зависимости
от уровня их обмена, — 35–85 мм рт. ст. Венозная кровь, поступающая
в легкие, имеет РО2 около 40 мм рт. ст.
Уже в альвеолярном воздухе содержание О2 и его парциальное
давление отличаются от атмосферного воздуха (13,6 % и 104 мм рт.
ст. соответственно). Это происходит вследствие нескольких причин:
- альвеолярный воздух лишь частично замещается атмосферным во время
каждого вдоха; - О2 постоянно абсорбируется из альвеолярного воздуха;
- СО2 постоянно диффундирует из крови легочных капилляров
в альвеолы; - сухой атмосферный воздух, который поступает в дыхательные пути, увлажняется,
насыщаясь водными парами, прежде чем достигает альвеол.
В артериальной крови, покидающей легкие, РО2
уменьшается уже до 95 мм рт. ст. вследствие так называемого венозного
примешивания. Дело в том, что определенное количество венозной крови
(1–2 %) не аэрируется в легких, поступая напрямую в артериальное
русло, что и приводит к некоторому снижению РО2.
Увеличение объема шунтированной крови, которое происходит при гипоксии
может приводить к существенному понижению артериального РО2.
Увеличение толщины альвеолокапиллярной мембраны, затрудняющее диффузию
газов, часто является следствием отека — увеличения количества жидкости
в межклеточном пространстве мембраны. Состояние характеризуется как
интерстициальный отек легких. Кроме того, жидкость может накапливаться
и в альвеолах, так что газам приходится проходить не только через
мембрану, но и через жидкость, что характерно уже для альвеолярного
отека легких. В большинстве случаев имеет место сочетание этих состояний
той или иной степени выраженности. Имеет значение и исходное состояние
легких. При ряде заболеваний, может возникнуть фиброз какого либо
участка легких с утолщением альвеолокапиллярной мембраны. Поскольку
скорость диффузии газов через мембрану обратно пропорциональна ее
толщине, любой фактор, увеличивающий ее толщину более чем в два –
три раза, может существенно нарушать нормальный газообмен.
Из капилляров кровеносного русла О2 диффундирует
в периферические ткани, поскольку парциальное давление О2
в капиллярах выше, чем в тканях. РО2 в интерстициальной
жидкости вне капилляров составляет в среднем 40 мм рт. ст., тогда
как в артериальной крови — 95 мм рт. ст. В венозной крови, покидающей
капилляр, РО2 также составляет около 40 мм рт. ст.
Если скорость тканевого кровотока увеличивается, большие количества
О2 доставляются тканям и тканевое РО2
увеличивается. Увеличение тканевого метаболизма приводит к снижению
РО2 как в периферических тканях, так и в венозной
крови.
Поскольку О2 постоянно используется тканями, внутриклеточное
РО2 всегда остается ниже интерстициального. Со снижением
интерстициального РО2 снижается и РО2
внутри клеток, повышение интерстициального РО2 ведет
к повышению внутриклеточного РО2. Важным общим свойством
живых организмов является большой запас функциональной прочности систем
поддержания жизнедеятельности. Такая особенность характерна и для
процессов газообмена. Для полного обеспечения процессов метаболизма
достаточно РО2 1–5 мм рт. ст., в то время как внутриклеточное
РО2 может варьировать от 5 до 60 мм рт. ст. (в зависимости
от протяженности капиллярного русла). Поэтому организм человека и
может функционировать при весьма существенном снижении РО2
в атмосферном воздухе.
В нормальных условиях около 97 % кислорода от легких к тканям переносится
в химически связанном виде гемоглобином. Лишь 3 % составляет О2
растворенный в плазме крови. Следует учитывать, что способность гемоглобина
связывать О2 является ограниченной. Каждый грамм
гемоглобина может максимально связать 1,34 мл О2.
Эта так называемая константа Гюффнера. Соответственно кислородная
емкость крови (т. е. максимальное общее количество кислорода, которое
может быть перенесено кровью) будет находиться в прямой зависимости
от содержания гемоглобина:
Кислородная емкость крови = [ Hb ]
x
1, 34 мл O2 / 100 мл крови
У здоровых людей с содержанием гемоглобина 150 г/л кислородная емкость
крови составляет 201 мл О2/л крови. Реально переносимое
количество мл О2 обычно меньше.
Ключевым фактором, характеризующим количество кислорода, связанного
с гемоглобином, является насыщение артериальной крови кислородом (сатурация,
SaO2). Оно выражает отношение между количеством кислорода,
связанного с гемоглобином и кислородной емкостью крови:
SaO2 = ( HbO2 / кислородная емкость крови )
x
100 %
Содержание кислорода в крови (контент, СаО2) — это
сумма связанного с гемоглобином и растворенного в плазме О2:
CaO2 = ( 1, 34
x
[ Hb ]
x
SaO2 ) + ( PaO2
x
0, 0031 )
Очевидно, что СаО2 зависит главным образом от фракции
О2, связанной с гемоглобином.
Кровь содержит незначительное количество кислорода, не связанного
с гемоглобином, а растворенного в плазме. Согласно закону Генри, количество
растворенного кислорода пропорционально парциальному давлению О2
и коэффициенту его растворимости, а растворимость О2
в крови очень низка: только 0,0031 мл О2 растворяется
в 0,1 л крови при увеличении давления на 1 мм рт. ст. Таким образом,
при РаО2 равным 100 мм рт. ст., в 100 мл артериальной
крови содержится только 0,31 мл растворенного О2.
Со снижением РаО2 количество растворенного в плазме
О2 станет еще меньше.
Содержание О2 в связи с изменениями РаО2
колеблется незначительно до тех пор, пока устойчиво поддерживается
SaO2. Изменения содержания гемоглобина приводят к
более заметным сдвигам СаО2. Нормальное СаО2
равно 198 мл О2/л крови при условии, что
PaO2 = 100 мм рт. ст.,
содержание гемоглобина 150 г/л, а
SaO2 = 97 %.
Умеренная анемия (например гемоглобин 120 г/л) при поддержании нормального
РаО2 проявляется снижением СаО2
до 160 мл О2/л крови.
Сродство гемоглобина к кислороду возрастает по мере последовательного
связывания молекул О2, что придает кривой диссоциации
оксигемоглобина сигмовидную или S-образную форму. Эта кривая, соотносящая
изменения SaO2 в зависимости от РаО2,
важна для анализа процессов транспорта кислорода к периферическим
тканям:
Верхняя часть кривой (при РаО2 > 60
мм рт. ст.) — относительно плоская. Это приводит к тому, что SaO2,
а следовательно, и СаО2 остаются достаточно постоянными,
несмотря на значительные колебания РаО2. Повышение
СаО2 или транспорта кислорода в этой области кривой
может быть достигнуто только за счет увеличения содержания гемоглобина
(например, при переливании крови) или растворения в плазме крови кислорода
(например при гипербарической оксигенации).
Крутые средняя и нижняя часть кривой иллюстрируют то положение, что,
хотя SaO2 падает (когда РаО2 оказывается
ниже 60 мм рт. ст.), процесс насыщения гемоглобина кислородом продолжается,
поскольку градиент РаО2 между альвеолами и капиллярами
сохраняется. Периферические ткани в этих условиях могут продолжать
извлекать достаточное количество О2, несмотря на
снижение капиллярного РО2.
РаО2, при котором гемоглобин насыщен кислородом на
50 % (при 37 °С и рН 7,4) известно как Р50. Это
общепринятая мера сродства гемоглобина к кислороду. Р50
в крови человека в норме составляет 26,6 мм. рт. ст. Однако оно может
изменяться при различных метаболических и физиологических условиях,
воздействующих на процесс связывания кислорода гемоглобином.
Когда сродство гемоглобина к кислороду падает, О2
с большой легкостью переходит в ткани, и наоборот. Повышение Р50
определяет сдвиг кривой диссоциации оксигемоглобина вправо. Сродство
гемоглобина к кислороду снижается, указывая, что теперь требуется
более высокое РаО2 для поддержания SaO2
на прежнем уровне. Более низкое сродство гемоглобина к кислороду означает
повышенное высвобождение кислорода в тканях, но ухудшение связывания
гемоглобина с О2 в легких.
Сдвиг кривой диссоциации влево и соответствующее снижение Р50
указывает на повышенное сродство гемоглобина к кислороду — улучшение
связывания в легких и ухудшение высвобождения О2
в периферических тканях. На Р50 и положение кривой
диссоциации гемоглобина влияет несколько факторов, в частности, рН
и температура.
Для обеспечения обмена веществ в периферических тканях важное значение
имеет не только общее содержание О2 — ключевой физиологической
переменной является его доставка. Доставка кислорода — это количество
кислорода, транспортируемое к тканям в единицу времени. Чтобы поддержать
аэробный метаболизм и предотвратить накопление лактата, периферические
ткани должны постоянно снабжаться кислородом. Обстоятельства, которые
определяют адекватность кислородного снабжения, разнообразны и включают
состояние покоя, физическую нагрузку, гиперкатаболические состояния
и инфекцию.
Доставка кислорода к периферическим тканям зависит от количества кислорода
в определенном объеме крови и уровня кровотока. Хотя объем кровотока
к отдельным органам различен, в периферических тканях он в целом равен
сердечному выбросу (CB). Системная доставка О2 (DО2)
рассчитывается как
DO2 = СВ (л/мин) x CaO2 (мл/л).
Нередко DО2 рассчитывают с учетом площади тела. То
есть на самом деле рассчитывают индекс DО2. Нормальными
величинами индекса DО2 считают 520–720 мл·мин-1·м-2.
Доставка кислорода падает при уменьшении сердечного выброса или снижении
объемного содержания кислорода в артериальной крови. Это характерно
для многих критических и терминальных состояний и чаще всего наблюдается
при выраженных проявлениях сердечной недостаточности, повышении внутрилегочного
шунтирования крови, централизации кровообращения, при различных пороках
сердца, кардиохирургических и других длительных и травматичных вмешательствах.
С практической точки зрения следует иметь в виду, что любое нарушение
кровообращения будет ухудшать доставку О2.
Потребление кислорода является заключительным этапом транспорта кислорода
тканям и представляет собой кислородное обеспечение тканевого метаболизма.
В условиях основного обмена взрослый человек потребляет около 250
мл О2 в 1 мин. Однако скорость утилизации О2
различными тканями значительно отличается.
Потребление кислорода тканями (VO2) — интегральный
показатель, учитывающий как циркуляторный компонент транспорта кислорода
(сердечный выброс), так и его гемический компонент (артериовенозное
различие по кислороду,
CaO2 — CvO2).
Его можно определить по формуле:
VO2 = СВ x (CaO2 — CvO2) x 10.
Так же как и DО2, VO2 часто выражают
в виде индекса, то есть в перерасчете на площадь тела. Нормальными
величинами индекса потребления кислорода являются 110–160 мл·мин-1·м-2.
В зависимости от состояния организма (покой, нагрузка или заболевание)
меняется фракционное распределение СВ к органам. Более того, и экстракция
кислорода тканями различных органов неодинакова. Например, миокард
получает лишь малую фракцию СВ, но извлекает почти весь доставляемый
кислород.
Нормальный компенсаторный ответ на снижение кровотока проявляется
в виде увеличения поглощения кислорода, достаточного для поддержания
VO2 на нормальном уровне. Падение сердечного выброса
компенсируется увеличением артериовенозного различия по кислороду,
и VO2 остается неизменным. Снижение насыщения венозной
крови кислородом отражает увеличение экстракции кислорода.
Способность компенсировать снижение кровотока повышением поглощения
кислорода является характерной особенностью микроциркуляторного русла
практически всех органов и тканей за исключением сердца и диафрагмы.
В них высокая экстракция кислорода из капиллярного ложа происходит
уже в норме. Поэтому уровень кислорода в тканях сердца и диафрагмы
весьма чувствителен даже к незначительным изменениям кровотока.
Таким образом, основными факторами, определяющими доставку О2,
являются:
- парциальное давление кислорода в атмосферном воздухе;
- нормальное состояние альвеолокапиллярной мембраны;
- эффективность дыхательной системы;
- достаточное количество гемоглобина крови;
- эффективность системы кровообращения.
Сергей Науменко,
г. Новосибирск, 2018 г.
Источник