Кровь у дождевых червей гемоглобин
Содержание статьи
Кровеносная система. Функции распределения питательных веществ и кислорода — дождевых червей
Распределение питательных веществ, всосанных поверхностью кишечника, осуществляется у дождевых червей при помощи высокоразвитой кровеносной системы. Схема расположения главных сосудов ее следующая (рисунок 12). Вдоль всего тела проходят спинной (над кишечником) и брюшной (под кишечником) сосуды. Спинной сосуд снабжен мускулатурой, которая волнообразными сокращениями гонит кровь от заднего конца тела к переднему. В нескольких передних сегментах (с 7-го по 11-й или, у других видов, с 7-го по 13-й) спинной сосуд сообщается 5 — 7 парами поперечных сосудов с брюшным сосудом. Эти сосуды снабжены особенно сильной мускулатурой и называются сердцами. Они вполне оправдывают это название, так как служат главным аппаратом, обеспечивающим кровообращение. Кровь, поступающая из сердец в брюшной сосуд, движется по направлению к заднему концу тела. По пути она поступает в сосуды, питающие стенку тела, а также в сосуды, идущие к кишечнику, к выделительным органам (рисунок 13), а в соответственных сегментах и к половым органам. Во всех этих частях тела сосуды распадаются на сеть микроскопических капилляров. Из капилляров кровь поступает в поперечные сосуды, которые собирают в конечном счете кровь из всего тела в спинной сосуд.
Имеются и другие продольные и-поперечные сосуды, которые можно видеть на рисунке 12 и 13; на них мы останавливаться не будем. Особое значение имеет густое сплетение мелких сосудов вокруг кишечника (рисунок 13). Сюда поступают питательные вещества, всосанные кишечником, и отсюда они разносятся по всему телу. Заметим, что мускулатурой, хотя и не столь сильно развитой, как в спинном сосуде и сердцах, обладают почти все сосуды, предотвращается возможность застоя крови в периферических частях кровеносной системы.
Кровь у дождевых червей, как уже было отмечено, красная. Этот ее цвет обусловлен наличием вещества, очень близкого к гемоглобину крови позвоночных.
Рисунок. 12. Дождевой червь, вскрытый со сплошной стороны. (Из Н. А. Холодковского).
Однако у червей содержится не в тельцах крови, а растворено в жидкой части крови (кровяной плазме). У дождевых червей имеются в крови только бесцветные клетки нескольких типов, в общем тех же, что и типы бесцветных клеток крови у позвоночных.
Рисунок. 13 Схема кровообращения в сегменте средней части тела дождевого червя на поперечном разрезе. (Из Штольте).
I — спинной сосуд; 2 — спинно-кишечный сосуд; 3 — кольцевой сосуд; 4 — сосуд тифлозоля; 6 — кишечное сплетение; 6 — сосуды нефридия; 7 — брюшной сосуд; 8 — сосуды брюшной нервной цепочки. Черной линией обозначено кишечное сплетение.
Как известно, гемоглобин у позвоночных обеспечивает транспорт кислорода из органов дыхания ко всем живым клеткам тела. Ту же роль играет вещество, сходное с гемоглобином, и у дождевых червей. Однако специальных дыхательных органов у них нет: они дышат всей поверхностью тела. Тонкая кутикула и нежность кожных покровов дождевых червей, а также богатая сеть кожных кровеносных сосудов, хорошо обеспечивают возможность поглощения кислорода из окружающей среды. Но отметим, что кутикула дождевых червей хорошо смачивается водой и кислород, по-видимому, у них должен сначала раствориться и йоде, смачивающей кожные покровы.
Это влечет за собой необходимость сохранения кожи во влажном состоянии. Уже это одно делает ясным, насколько важными для жизни червей являются условия влажности внешней среды. С подсыханием кожи дыхание для них делается невозможным. Однако при попадании в условия недостатка влаги в почве червь может долгое время бороться с этим, используя запасы воды, имеющиеся внутри тела. В этих случаях ему на помощь приходят кожные железы (смотрите страница 15), а в случае острого недостатка влаги он утилизирует для этого полостную жидкость, выбрызгивая ее из спинных пор.
Рисунок. 14. Капилляры в кожном эпителии дождевого червя. (Из Штольте).
1 — аиидермио; 2 — кольцевал мускулатура; 3 — продольная мускулатура.
Поглощение кислорода поверхностью тела облегчается наличием очень богатой сети кровеносных капилляров, проникающих даже в кожный эпителий (рисунок 14) Отсюда кровь через сосуды стенки тела и поперечные сосуды попадает в главные стволы кровяного русла, чем достигается снабжение кислородом всего тела. Красные топа окраски большинства видов дождевых червей определяются именно, наличием богатой сети кожных кровеносных сосудов.
Все это создает возможность жизни червей в условиях очень пониженного содержания кислорода.
В этом отношении они приближаются к некоторым из своих отдаленных пресноводных родичей — червям-трубочникам (Ти- bifex tubifex, Limnodrilus hoffmeisteri и др.), которые, обитая в глубоком илу, могут выносить почти полное отсутствие кислорода. Относительно дождевых червей есть наблюдения, что они могут жить при содержании кислорода в окружающем их воздушном пространстве, равном 2.5% (как известно, обычно в воздухе его содержится 21/о). Даже при наличии 0.4% кислорода в воздухе черви могут поглотить половину количества кислорода, необходимого им для поддержания жизни, и могут находиться в этих условиях довольно долго. Кроме того, попав в бескислородную среду, черви могут переключаться на особый тип обмена веществ, в котором источником энергии для жизненных процессов служат не реакции окисления (для которых и нужен кислород), а распад крахмало подобного вещества — гликогена, который совершается и в бескислородной среде. Однако запасы гликогена у червей не особенно велики, и, кроме того, при этом способе обмена веществ выделяются кислоты, вредно действующие на организм червей.
Находясь под водой, черви столь же хорошо могут поглощать кислород, как и в воздушной среде. Известно, что они могут жить в воде месяцами при наличии нужного им минимума кислорода н других необходимых для них условий. Этот факт имеет большое значение для понимания многих явлений из жизни дождевых червей.
1 Это вещество получило название «гемокруорнн».
Также рекомендуем прочитать:
- Eisenia veneta — Венецианская — вид дождевых червей
- Враги и паразиты — дождевых червей
- Выращивание дождевых червей для корма цыплят. (способ М. М. Исаковой — Кео)
- Кишечник и питание — дождевых червей
- Методы вскрытия — дождевых червей
- Мускулатура и движение. Полость телa — дождевых червей
- Нервная система и органы чувств. Рефлексы — дождевых червей
- Органы внутренней секреции — дождевых червей
- Органы выделения. Поглощение и выделение воды — дождевых червей
- Органы размножения — дождевых червей
- Польза и вред — дождевых червей
- Способы сбора и консервированиия — дождевых червей
- Строение тела и главные жизненные отправления дождевых червей — общие замечания. наружные признаки
- Влияние внешних условий на интенсивность образования выбросов дождевых червей на поверхность почвы
- Влияние дождевых червей на перемешивание земли и вынос ее на поверхность
Источник
ождевой червь. Класс Oligochaeta. Строение дождевого червя. Размножение дождевого червя.
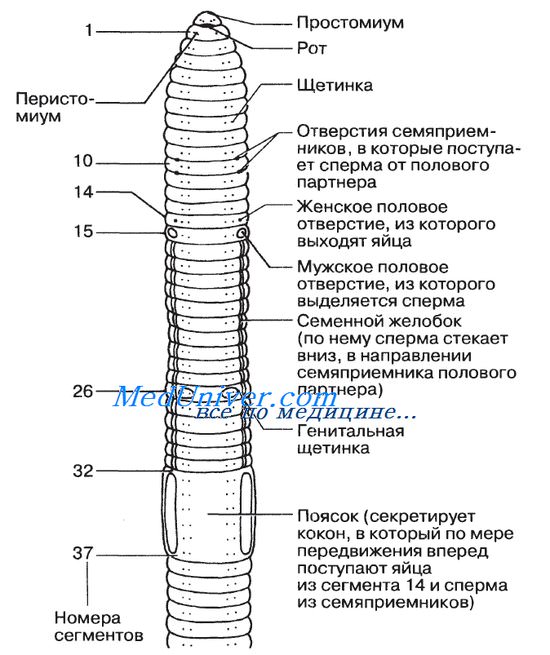
Оглавление темы «Членистоногие. Хордовые.»: Дождевой червь. Класс Oligochaeta. Строение дождевого червя. Размножение дождевого червя.Дождевой червь (Lumbricus terrestris) — животное с длинным цилиндрическим телом, достигающее в длину приблизительно 12—18 см. Передний его конец конусообразный, а задний сплющен в дорсовентральном направлении. И хотя дождевой червь живет на суше, он не смог до конца преодолеть все проблемы, связанные с наземным образом жизни. Спасаясь от высыхания, он живет под землей, прорывая нору во влажной почве, и покидает ее лишь по ночам, отправляясь на поиски пищи или полового партнера. Различия во внешнем строении тела Nereis и дождевого червя обусловлены приспособлением последнего к наземному образу жизни. Тело дождевого червя имеет обтекаемую форму и лишено каких-либо выростов, наличие которых могло бы препятствовать свободному передвижению червя в почве. Простомиум, расположенный над ртом, небольшой и округлый, чувствительных придатков на нем нет. Все сегменты, за исключением первого и последнего, несут по четыре пары щетинок; две из них расположены вент-рально (на брюшной стороне), а две дорсо-латерально. Щетинки дождевого червя выходят из расположенных в стенке тела щитинковых мешков. Они могут втягиваться внутрь или выводиться наружу под действием специальных мышц. Щетинки облегчают передвижение червя по земле. Щетинки используются также и для защиты: цепляясь ими за стенки норы, он прочно удерживается в ней. Самые длинные щетинки, расположенные на сегментах 10—15, 26 и 32—37, используются при копуляции. Еще одна структура, используемая при копуляции, — поясок, расположенный на сегментах 32—37. Эпидермис пояска содержит множество железистых клеток, образующих на спинной и боковых поверхностях некоторое утолщение, напоминающее по форме седло. Поясок участвует в процессах копуляции и формирования кокона.
Ротовое и анальное отверстия дождевого червя расположены на противоположных концах тела. Дождевой червь питается детритом (фрагментами разлагающегося органического материала), который червь заглатывает вместе с почвой. Подавляющая часть поглощенной почвы проходит через кишечник и затем выбрасывается на поверхность почвы в виде характерных кучек. Выделяющаяся через спинные поры дождевого червя целомическая жидкость и слизь, секретируемая железистыми клетками эпидермиса, постоянно увлажняют тонкую кутикулу. В результате предотвращается высыхание кутикулы, облегчается газообмен и создается своего рода смазка, облегчающая передвижение червя в почве. Именно через кутикулу происходит газообмен путем диффузии — процесс, зависимый от наличия в эпидермальном слое разветвленной сети капилляров. Все сегменты дождевого червя, за исключением трех первых и одного последнего, несут по паре нефридиев — канальцев, выполняющих функции выделения и осморегуляции. Они открываются на поверхности тела порами. Репродуктивная система дождевого червя, как и поведение, связанное с размножением, отличается большой сложностью, что, возможно, объясняется наземным образом жизни и необходимостью предохранять от высыхания гаметы и оплодотворенные яйца. Lumbricus гермафродит (т. е. у каждой особи имеются и мужские и женские репродуктивные органы). Контакт дождевых червей друг с другом происходит очень редко, но когда это происходит, спариваться могут две любые особи, поскольку обе они гермафродиты. В результате происходит взаимообмен спермой и оба червя оплодотворяются. Половые органы дождевых червей сосредоточены на переднем конце тела. Внешние признаки, связанные с половым размножением, приведены на рисунке. Спаривание и последующая откладка оплодотворенных яиц — процесс, очень сложный. Спаривающиеся особи располагаются рядом друг с другом во взаимно противоположных направлениях, так что голова одного из партнеров оказывается направленной в сторону хвостового отдела другого и наоборот. — Также рекомендуем «Членистоногие. Тип Arthropoda. Строение членистоногих. Экзоскелет. Кутикула.» |
Источник
ыхание дождевого червя. Дыхательная система насекомых
Дыхание дождевого червя. Дыхательная система насекомых — саранчи.Дождевой червь относится к группе кольчатых червей. У него нет никаких особых органов, предназначенных специально для газообмена, и газообмен происходит путем диффузии через всю поверхность тела. Специализированные органы им в сущности и не нужны, так как благодаря цилиндрической форме тела отношение площади поверхности к объему у них велико, а при своей относительно малой активности они расходуют не так много кислорода. Однако у кольчатых червей имеется кровеносная система (в отличие от некоторых более простых животных и одноклеточных организмов), а в их крови растворен дыхательный пигмент гемоглобин. Сокращения крупных кровеносных сосудов прогоняют кровь вместе с растворенными в ней газами по всему телу; это же способствует и поддержанию крутых диффузионных градиентов. Тонкая кожа дождевого червя (кутикула) постоянно увлажняется секретом находящихся в эпителии желез. В эпителии непосредственно под кутикулой расположены капилляры. Расстояние между кровеносными сосудами и поверхностью тела невелико и это обеспечивает быструю диффузию кислорода в кровь. Дождевые черви практически ничем не защищены от высыхания и поэтому стараются держаться только во влажной среде.
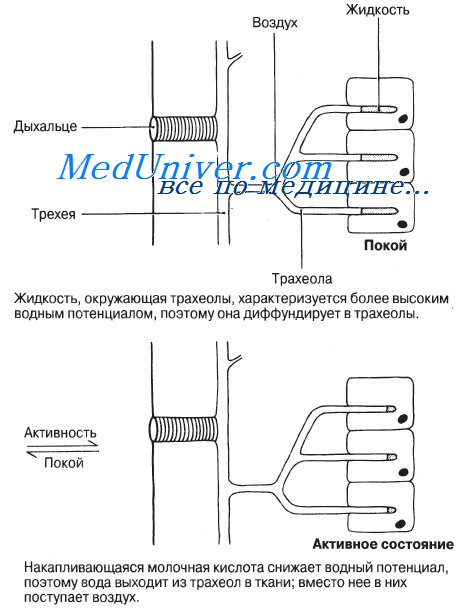
Дыхательная система насекомых — саранчи.У насекомых газообмен осуществляется через систему трубочек, так называемых трахей. Такая система позволяет кислороду поступать из воздуха прямо к тканям и необходимость в его транспортировке кровью отпадает. Это гораздо более быстрый способ, нежели диффузия растворенного кислорода сквозь ткани; такой газообмен создает условия для высокой интенсивности метаболизма. Дыхальца — парные отверстия, имеющиеся на втором и третьем грудном и на первых восьми брюшных сегментах тела насекомого, ведут в воздушные полости. От этих полостей отходят разветвленные трубочки — трахеи. Каждая трахея выстлана эпителием, секретирующим тонкий слой хитинового материала. Обычно этот жесткий слой еще более укреплен спиральными и кольцевыми утолщениями, благодаря которым воздухоносные пути остаются открытыми, даже если в просвете трахей давление оказывается отрицательным (сравните с хрящевыми кольцами в трахее и бронхах человека). В каждом сегменте тела трахеи разветвляются на многочисленные более мелкие трубочки, называемые трахеолами; трахеолы тоже ветвятся, пронизывая ткани насекомого, и в наиболее активных тканях, например в летательных мышцах, оканчиваются слепо внутри отдельных клеток. Степень ветвления трахеол может меняться в зависимости от метаболических нужд тканей. В трахеолах хитиновая выстилка отсутствует. В состоянии покоя они наполнены водянистой жидкостью; в это время кислород диффундирует по ним к тканям (а С02 — в обратном направлении) со скоростью, вполне достаточной для удовлетворения потребностей насекомого. В активном состоянии усиление метаболической активности мышц ведет к накоплению определенных метаболитов, в частности молочной кислоты, и в тканях соответственно повышается осмотическое давление. Когда это происходит, жидкость из трахеол под действием осмотических сил частично всасывается в ткани, и в трахеолы поступает больше воздуха, а значит, и больше кислорода, причем этот кислород подается непосредственно к тканям как раз тогда, когда они в нем нуждаются.
Общий поток воздуха, проходящий через тело насекомого, регулируется механизмом, закрывающим дыхальца. Отверстие каждого дыхальца снабжено системой клапанов, управляемых очень мелкими мышцами. Края этого отверстия покрыты волосками, которые препятствуют попаданию в дыхальца чужеродных частиц и предотвращают излишнюю потерю влаги. Величина отверстия регулируется в зависимости от количества С02 в теле насекомого. Усиленная активность ведет к усиленному образованию СО2. Хеморецепторы улавливают это и дыхальца открываются. Тот же стимул может вызывать и вентиляционные движения тела, особенно у крупных насекомых, таких как саранча. Дорсовентральные мышцы, сокращаясь, делают тело насекомого более плоским, вследствие чего объем трахейной системы уменьшается и воздух выталкивается из нее наружу («выдох»). Всасывание воздуха («вдох») происходит пассивно, когда сегменты тела благодаря своей эластичности принимают исходную форму. Судя по некоторым данным, грудные и брюшные дыхальца открываются и закрываются попеременно, и это в сочетании с вентиляционными движениями тела создает однонаправленный поток воздуха, который входит в тело насекомого через грудной отдел и выходит через брюшной. Трахейная система, безусловно, весьма эффективна в смысле газообмена, однако следует учитывать, что газообмен определяется у большинства насекомых исключительно диффузией кислорода через ткани насекомого. Диффузия же, как известно, эффективна только на малых расстояниях, и это накладывает жесткие ограничения на размеры, которых могут достигать насекомые. Эти малые расстояния, на которых диффузия достаточно эффективна, не превышают 1 см; поэтому, хотя и встречаются насекомые длиной до 30 см, их тело не должно при этом иметь в толщину более 2 см. — Также рекомендуем «Дыхание костных рыб — сельди.» Оглавление темы «Вентиляция легких.»: |

Источник
Эволюционный дауншифтинг дождевых червей
Представьте, что самый обычный дождевой червяк стал длинным и толстым, как шлагбаум, и начал жить в трубке, которую строит сам и откуда торчат лишь его многочисленные щупальца. Из своей трубки он никогда не вылезает и селится на останках костей позвоночных, гниющих обломках затонувших судов или близ сероводородных источников. Сложновато вообразить такого уже далеко не дождевого червя, не правда ли? Но в природе такие организмы существуют, а имя им — вестиментиферы.

Вестиментифера Riftia pachyptila без трубки
Питер Бэтсон
Черви-вестиментиферы вместе со своими сородичами погонофорами, костоедами и древоточцами относятся к семейству зибоглинид (понимаем, имена у этих червей на любителя). Все эти группы червей относятся к сидячим кольчатым червям и произошли от того же предка, что и дождевой червь. Только эволюция загнала дождевого червя в почву, а зибоглинид отправила осваивать глубоководные местообитания. Как мы уже упоминали, некоторые вестиментиферы селятся близ гидротермальных источников, а условия там далеко не райские для нормальных аэробных организмов. В общем, вестиментифер можно считать примером эволюционного дауншифтинга среди червяков.
Расскажем немножко подробнее про обстановку близ гидротермальных источников, которые считаются колыбелью жизни на Земле. Температура там зависит от активности источника и может колебаться в диапазоне от +2 до +30 градусов Цельсия, реакция среды кислая (pH может опускаться до 4,4), химический состав сильно варьирует, концентрация сероводорода и сульфидов превышает допустимую для человека норму в 100 раз… То холодно, то жарко, кисло и крайне токсично для большинства аэробных организмов (сероводород блокирует конечный фермент дыхательной цепи). Но наши герои, черви-вестиментиферы, сумели успешно приспособиться к ссылке на сероводородные источники и даже извлекают пользу из ядовитового окружения.
Отметим, что далеко не все вестиментиферы селятся близ гидротермальных источников, к ним тяготеют лишь некоторые представители, такие как рифтия (Riftia pachyptila) и риджея (Ridgeia piscesae). Эти черви избавились от кишечника, и почти весь просвет их целома занимает специальный орган — трофосома. Прямо внутри клеток этого органа живут симбиотические хемоавтотрофные бактерии. Такие бактерии используют гидросульфид-ион (HS-) в качестве источника окисления, освобождающиеся в ходе окисления электроны поступают в электрон-транспортную цепь, и в конечном счете образуется АТФ — универсальная энергетическая валюта живых существ (по крайней мере, на Земле).
Все питание наших вестиментифер построено на симбиотических отношениях с хемоавтотрофными бактериями: черви снабжают симбионтов жилплощадью, обеспечивают сульфидом, а взамен получают готовую органику в виде сукцината и глутамата. Встает вопрос: как транспортировать через свое тело токсичные сульфиды, чтобы «накормить» бактерий? На этот случай у рифтии и риджеи есть измененный гемоглобин. Цепочки фермента у этих червей снабжены цистеиновыми остатками, способными связываться с сероводородом и транспортировать его по телу (с переносом кислорода при этом у них проблем не возникает).

Электронограмма, сделанная на трансмиссионном электронном микроскопе, бактерии трофосомы рифтии
Надежда Римская-Корсакова
Кроме того, недавно мы на кафедре зоологии беспозвоночных животных биофака МГУ исследовали кровеносную систему рифтии, гиганта-рекордсменки среди вестиментифер, и выявили ряд характерных особенностей, не свойственных прочим кольчатым червям. У рифтии оказалась хорошо развита лакунарная система, дренирующая основные системы органов (в том числе трофосому, где живут бактерии-симбионты), основные сосуды, по которым проталкивается кровь к верхнему концу тела червя, дополнительно выстланы слоем мышечной тканью.
Кроме того, в щупальцевой зоне рифтии соединяются две системы сосудов, из-за чего кровь к щупальцам идет как по базальным, так и по аксиальным сосудам. Дополнительная сосудистая система позволила рифтии обзавестись аж 400 парами ламелл, несущих щупальца (у других вестиментифер их число не превышает 70).

Схема кровоснабжения вестиментиферы рифтии
Надежда Римская-Корсакова
Таким образом, измененный гемоглобин и усиленная кровеносная система позволили родне дождевых червей стать резистентными к сильному токсину, перейти целиком на симбиотическое питание и освоить малопригодную для жизни нишу.
Литература
Tunnicliffe, V., Germain, C. S., & Hilario, A. Phenotypic variation and fitness in a metapopulation of tubeworms (Ridgeia piscesae Jones) at hydrothermal vents // PloS one, 2014. — 9(10).
Bright, M., and F. H. Lallier. The biology of vestimentiferan tubeworms // Oceanogr. Mar. Biol. Annu. Rev., 2010 (48). — Pp. 211–264.
Bailly, X., & Vinogradov, S. The sulfide binding function of annelid hemoglobins: relic of an old biosystem? // Journal of inorganic biochemistry, 2005. — 99(1). — Pp. 142-150.
Rimskaya-Korsakova, N. N., Galkin, S. V., & Malakhov, V. V. The anatomy of the blood vascular system of the giant vestimentiferan tubeworm Riftia pachyptila (Siboglinidae, Annelida) // Journal of Morphology, 2017. — 278(6). — Pp. 810-827.
Надежда Римская-Корсакова,
кандидат биологических наук,
ведущий научный сотрудник кафедры зоологии беспозвоночных биологического факультета МГУ имени М.В. Ломоносова
Источник