Производные холестерина в организме
Содержание статьи
18. Биологически активные производные холестерина.
К производным холестерина относятся стероидные гормоны и жёлчные кислоты.
Холестерин, который находится в организме человека, подвержен окислению. Так он преобразуется во всевозможные стероидные соединения. Примерно 70% имеющегося в организме свободного холестерина уходит на это процесс. Желчные кислоты синтезируются в печени, а затем хранятся и концентрируются в желчном пузыре. При необходимости они направляются в тонкий кишечник. Самой значимой в организме является холевая кислота. Еще есть дезоксихолевая, хенодезоксихолевая и литохолевая. Некоторые из них присутствуют в желчи в виде солей. Эти производные холестерина являются основными компонентами желчи. В них растворяются диетические липиды. Кроме того и сами они являются конечными продуктами распада этого вещества.
Производными холестерина являются 5 основных классов стероидных гормонов: прогестины, глюкокортикоиды, минералокортикоиды, андрогены и эстрогены. Эти гормоны являются мощными веществами, регулирующими основные функции в организме. Прогестерон и прогестаген подготавливает матку к имплантации яйцеклетки. Для нормального протекания беременности необходим прогестерон. Андрогены определяют вторичные мужские половые признаки, а эстрогены — женские. Глюкокортикоиды участвуют в образовании гликогена, подавляют воспалительные реакции. Минералокортикоиды воздействуют на почки, что приводит к увеличению потока крови и артериального давления.
Холестерин является предшественником витамина D, который играет важную роль в контроле метаболизма кальция и фосфора.
Вопрос 21 Биологическая роль вторичных мессенджеров при передаче гормонального сигнала
Общим фундаментальным механизмом, посредством которого реализуются биологические эффекты «вторичных» мессенджеров внутри клетки, является процесс фосфорилирования — дефосфорилирования белков при участии широкого разнообразия протеинкиназ, катализирующих транспорт концевой группы от АТФ на ОН-группы серина итреонина, а в ряде случаев — тирозина белков-мишеней. Процесс фосфорилирования представляет собой важнейшую посттрансляционную химическую модификацию белковых молекул, коренным образом изменяющую как их структуру, так и функции. В частности, он вызывает изменение структурных свойств (ассоциацию или диссоциациюсоставляющих субъединиц), активирование или ингибирование их каталитических свойств, в конечном итоге определяя скорость химических реакций и в целом функциональную активность клеток.
22. Стероидные гормоны. Механизм и бла бла бла
В отличие от пептидных, стероидные гормоны легко проникают через плазматическую мембрану клеток и взаимодействуют со своими рецепторами в цитоплазме и/или ядре клетки-мишени. Некоторые рецепторы стероидных гормонов — онкопротеины (например, erbA). Все рецепторы стероидных гормонов имеют ДНК-связывающий участок. Другими словами, рецепторы стероидных гормонов — факторы транскрипции. Конечный эффект взаимодействия стероидного гормона и его рецептора состоит в изменении спектра транскрибируемых генов. Таким образом, результат действия стероидных гормонов на клетку-мишень — индукция синтеза конкретных белков, что фундаментально изменяет метаболизм как клетки-мишени, так и множества других клеток организма. Синтезируемые под влиянием стероидных гормонов белки могут и сами быть гормонами или другими важными для функционирования клетки молекулами, например ферментами. После высвобождения из эндокринной клетки стероидные гормоны попадают в кровь, где около 95% гормонов связывается со специфическими транспортными белками (транскортинами, связывающими тестостерон белками, различными альбуминами и глобулинами). Рецепторы стероидных гормонов причисляют к обширной группе ядерных рецепторов, куда также относят рецепторы ретиноидов, витамина D3, трийодтиронина. После поступления в клетки-мишени молекулы стероидного гормона могут вызвать ответ только в том случае, если в клетке есть специфические внутриклеточные рецепторы для этого гормона. Так, эстрогеновые рецепторы обнаруживают в клетках-мишенях матки, молочной железы и мозга. Клетки волосяных фолликулов кожи лица и эректильная ткань полового члена содержат андрогеновые рецепторы. Глюкокортикоидные рецепторы обнаруживают практически во всех клетках. В клетке-мишени каждый из основных классов половых стероидных гормонов (андрогены, эстрогены, прогестины) индуцирует развитие цепи событий, которая включает (I) связывание стероида с его рецептором; (И) аллостерические конформационные изменения структуры рецептора, переводящие рецептор из неактивной формы в активную; (III) связывание стероид-рецепторного комплекса с регуляторными элементами ДНК; (IV) транскрипцию и синтез новых молекул м-РНК; (V) трансляцию м-РНК и синтез новых белков. При транскрипции РНК-полимераза II присоединяется к промотору — специфическому сайту молекулы ДНК, с которого начинается синтез полимера. РНК-полимераза II раскручивает участок двойной спирали ДНК, обнажая матрицу для комплементарного спаривания оснований. Когда РНК-полимераза встречает сигнал терминации транскрипции, синтез полимера прекращается. Большинство фармакологических и физиологических знаний о механизме действия стероидных гормонов было получено на основании исследований стероидных рецепторов. Эффективность действия стероидных гормонов зависит от аффинности рецептора для гормона или его фармакологического аналога, а также от эффективности аллостерически активированного комплекса гормон-рецептор в регуляции транскрипции.
23. механизм действия белковых гормонов….
Механизмы действия пептидных, белковых гормонов и катехоламинов. Лиганд. Молекулу гормона обычно называют первичным посредником регуляторного эффекта, или лигандом. Молекулы большинства гормонов связываются со специфическими для них рецепторами плазматических мембран клеток мишеней, образуя лиганд-рецепторный комплекс. Для пептидных, белковых гормонов и катехоламинов его образование является основным начальным звеном механизма действия и приводит к активации мембранных ферментов и образованию различных вторичных посредников гормонального регуляторного эффекта, реализующих свое действие в цитоплазме, органоидах и ядре клетки. Среди ферментов, активируемых лиганд-рецептор-ным комплексом, описаны: аденилатциклаза, гуанилатциклаза, фосфолипа-зы С, D и А2, тирозинкиназы, фосфаттирозинфосфатазы, фосфоинозитид-3-ОН-киназа, серинтреонин-киназа, синтаза N0 и др. Вторичными посредниками, образующимися под влиянием этих мембранных ферментов, являются: 1) циклический аденозинмонофосфат (цАМФ); 2) циклический гуано зинмонофосфат (цГМФ); 3) инозитол-3-фосфат (ИФЗ); 4) диацилглицерол; 5) олиго (А) (2,5-олигоизоаденилат); 6) Са2+ {ионизированный кальций); 7) фосфатидная кислота; 8) циклическая аденозиндифосфатрибоза; 9) N0 (оксид азота). Многие гормоны, образуя лиганд-рецепторные комплексы, вызывают активацию одновременно нескольких мембранных ферментов и, соответственно, вторичных посредников. Значительная часть гормонов и биологически активных веществ взаимодействуют с семейством рецепторов, связанных с G-белками плазматической мембраны (андреналин, норадреналин, аденозин, ангиотензин, эндотелии и др.).
Вопрос №26
Биохимическая роль нуклеотидов в метаболизме
Нуклеотиды — фосфорные эфиры нуклеозидов, нуклеозидфосфаты. Свободные нуклеотиды, в частности АТФ, цАМФ, АДФ, играют важную роль в энергетических и информационных внутриклеточных процессах, а также являются составляющими частями нуклеиновых кислот и многих коферментов. Биохимическая роль нуклеотидов:
Универсальный источник энергии (АТФ и его аналоги).
Являются активаторами и переносчиками мономеров в клетке(УДФ-глюкоза)
Выступают в роли коферментов (ФАД, ФМН, НАД+, НАДФ+)
Циклические мононуклеотиды являются вторичными посредниками при действии гормонов и других сигналов(цАМФ, цГМФ).
Аллостерические регуляторы активности ферментов.
Являются мономерами в составе нуклеиновых кислот, связанные 3′-5′- фосфодиэфирными связями.
Вопрос №27
Отличия и сходства строения ДНК и РНК
Дезоксирибонуклеиновая кислота (ДНК) — макромолекула (одна из трёх основных, две другие — РНК и белки), обеспечивающая хранение, передачу из поколения в поколение и реализацию генетической программы развития и функционирования живых организмов. ДНК содержит информацию о структуре различных видов РНК и белков.
С химической точки зрения ДНК — это длинная полимерная молекула, состоящая из повторяющихся блоков — нуклеотидов. Каждый нуклеотид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы. Связи между нуклеотидами в цепи образуются за счёт дезоксирибозы и фосфатной группы (фосфодиэфирные связи). В подавляющем большинстве случаев (кроме некоторых вирусов, содержащих одноцепочечную ДНК) макромолекула ДНК состоит из двух цепей, ориентированных азотистыми основаниями друг к другу. Эта двухцепочечная молекула спирализована. В целом структура молекулы ДНК получила название «двойной спирали».
В ДНК встречается четыре вида азотистых оснований (аденин, гуанин, тимин и цитозин). Азотистые основания одной из цепей соединены с азотистыми основаниями другой цепи водородными связями согласно принципу комплементарности: аденин соединяется только с тимином, гуанин — только с цитозином. Последовательность нуклеотидов позволяет «кодировать» информацию о различных типах РНК, наиболее важными из которых являются информационные, или матричные (мРНК), рибосомальные (рРНК) и транспортные (тРНК). Все эти типы РНК синтезируются на матрице ДНК за счёт копирования последовательности ДНК в последовательность РНК, синтезируемой в процессе транскрипции, и принимают участие в биосинтезе белков (процессе трансляции). Помимо кодирующих последовательностей, ДНК клеток содержит последовательности, выполняющие регуляторные и структурные функции.
Рибонуклеиновая кислота́ (РНК) — одна из трёх основных макромолекул (две другие — ДНК и белки), которые содержатся в клетках всех живых организмов.
Так же, как ДНК (дезоксирибонуклеиновая кислота), РНК состоит из длинной цепи, в которой каждое звено называется нуклеотидом. Каждый нуклеотид состоит из азотистого основания, сахара рибозы и фосфатной группы. Последовательность нуклеотидов позволяет РНК кодировать генетическую информацию. Все клеточные организмы используют РНК (мРНК) для программирования синтеза белков.
Нуклеотиды РНК состоят из сахара — рибозы, к которой в положении 1′ присоединено одно из оснований: аденин, гуанин, цитозин или урацил. Фосфатная группа соединяет рибозы в цепочку, образуя связи с 3′ атомом углерода одной рибозы и в 5′ положении другой. Фосфатные группы при физиологическом рН отрицательно заряжены, поэтому РНК — полианион. РНК транскрибируется как полимер четырёх оснований (аденина (A), гуанина (G), урацила (U) и цитозина (C), но в «зрелой» РНК есть много модифицированных оснований и сахаров). Всего в РНК насчитывается около 100 разных видов модифицированных нуклеотидов.
Азотистые основания в составе РНК могут образовывать водородные связи между цитозином и гуанином, аденином и урацилом, а также между гуанином и урацилом. Однако возможны и другие взаимодействия, например, несколько аденинов могут образовывать петлю, или петля, состоящая из четырёх нуклеотидов, в которой есть пара оснований аденин — гуанин.
Важная структурная особенность РНК, отличающая её от ДНК — наличие гидроксильной группы в 2′ положении рибозы, которая позволяет молекуле РНК существовать в А, а не В-конформации, наиболее часто наблюдаемой у ДНК. У А-формы глубокая и узкая большая бороздка и неглубокая и широкая малая бороздка. Второе последствие наличия 2′ гидроксильной группы состоит в том, что конформационно пластичные, то есть не принимающие участие в образовании двойной спирали, участки молекулы РНК могут химически атаковать другие фосфатные связи и их расщеплять.
«Рабочая» форма одноцепочечной молекулы РНК, как и у белков, часто обладает третичной структурой. Третичная структура образуется на основе элементов вторичной структуры, образуемой с помощью водородных связей внутри одной молекулы. Различают несколько типов элементов вторичной структуры — стебель-петли, петли и псевдоузлы.
Между ДНК и РНК есть три основных отличия:
ДНК содержит сахар дезоксирибозу, РНК — рибозу, у которой есть дополнительная, по сравнению с дезоксирибозой, гидроксильная группа. Эта группа увеличивает вероятность гидролиза молекулы, то есть уменьшает стабильность молекулы РНК.
Нуклеотид, комплементарный аденину, в РНК не тимин, как в ДНК, а урацил — неметилированная форма тимина.
ДНК существует в форме двойной спирали, состоящей из двух отдельных молекул. Молекулы РНК, в среднем, гораздо короче и преимущественно одноцепочечные.
Структурный анализ биологически активных молекул РНК, включая тРНК, рРНК, мяРНК и другие молекулы, которые не кодируют белков, показал, что они состоят не из одной длинной спирали, а из многочисленных коротких спиралей, расположенных близко друг к другу и образующих нечто, похожее на третичную структуру белка. В результате этого РНК может катализировать химические реакции, например, пептидил-трансферазный центр рибосомы, участвующий в образовании пептидной связи белков, полностью состоит из РНК.
Соседние файлы в предмете Биохимия
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник
Хороший, плохой, злой холестерин
Статья на конкурс «био/мол/текст»: Вряд ли сейчас найдется человек, который не слышал, что высокий холестерин — это плохо. Однако столь же мала вероятность встретить человека, который знает, ПОЧЕМУ высокий холестерин — это плохо. И чем определяется высокий холестерин. И что такое высокий холестерин. И что такое холестерин вообще, зачем он нужен и откуда берется.
Итак, история вопроса такова. Давным-давно, в одна тысяча девятьсот тринадцатом году, петербуржский физиолог Аничков Николай Александрович показал: не что иное, как холестерин, вызывает атеросклероз у экспериментальных кроликов, содержащихся на пище животного происхождения [1]. Вообще же, холестерин необходим для нормального функционирования животных клеток и является основной составляющей клеточных мембран , а также служит субстратом для синтеза стероидных гормонов и желчных кислот.
Главный липидный компонент пищевого жира и жировых отложений — это триглицериды, которые представляют собой эфиры глицерина и жирных кислот. Холестерин и триглицериды, будучи неполярными липидными веществами, транспортируются в плазме крови в составе липопротеиновых частиц. Частицы эти подразделяют по размеру, плотности, относительному содержанию холестерина, триглицеридов и белков на пять больших классов: хиломикроны, липопротеины очень низкой плотности (ЛПОНП), липопротеины промежуточной плотности (ЛППП), липопротеины низкой плотности (ЛПНП) и липопротеины высокой плотности (ЛПВП) [2]. Традиционно ЛПНП считается «плохим» холестерином, а ЛПВП — «хорошим» (рис. 1).
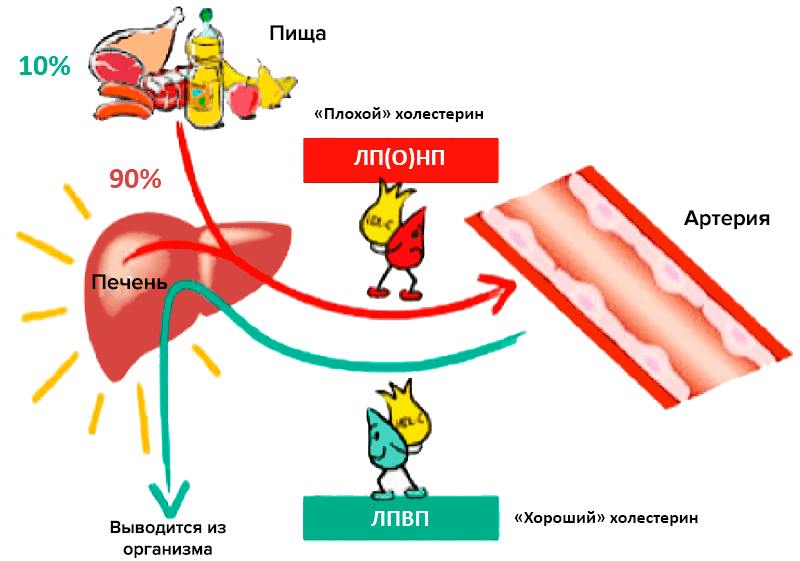
Рисунок 1. «Плохой» и «хороший» холестерины. Участие различных липопротеиновых частиц в транспорте липидов и холестерина.
Схематично структура липопротеина включает неполярное ядро, состоящее по большей части из холестерина и триглицеридов, и оболочку из фосфолипидов и апопротеинов (рис. 2). Ядро — функциональный груз, который доставляется до места назначения. Оболочка же участвует в распознавании клеточными рецепторами липопротеиновых частиц, а также в обмене липидными частями между различными липопротеинами [3].
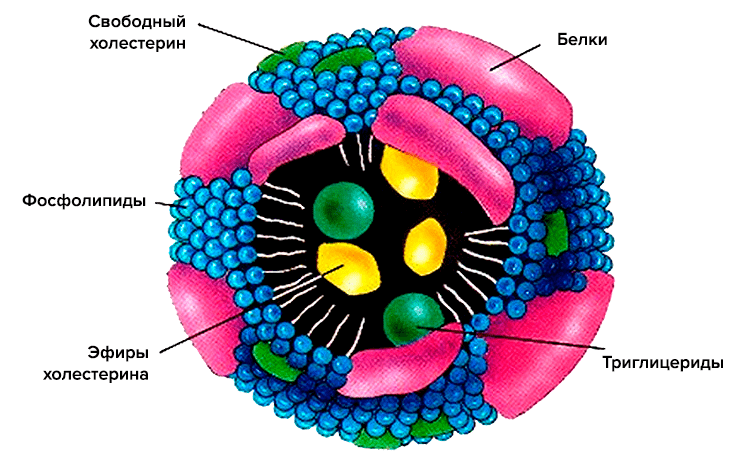
Рисунок 2. Схематическое строение липопротеиновой частицы
Баланс уровня холестерина в организме достигается следующими процессами: внутриклеточный синтез, захват из плазмы (главным образом из ЛПНП), выход из клетки в плазму (главным образом в составе ЛПВП). Предшественник синтеза стероидов — ацетил коэнзим А (CoA). Процесс синтеза включает, по крайней мере, 21 шаг, начиная с последовательного преобразования ацетоацетил CoA. Лимитирующая стадия синтеза холестерина в большой степени определяется количеством холестерина, абсорбируемого в кишечнике и транспортируемого в печень [4]. При недостатке холестерина происходит компенсаторное усиление его захвата и синтеза.
Транспорт холестерина
Систему транспорта липидов можно разделить на две большие части: внешнюю и внутреннюю.
Внешний путь начинается с всасывания в кишечнике холестерина и триглицеридов. Его конечный результат — доставка триглицеридов в жировую ткань и мышцы, а холестерина — в печень. В кишечнике пищевой холестерин и триглицериды связываются с апопротеинами и фосфолипидами, формируя хиломикроны, которые через лимфоток попадают в плазму, мышечную и жировую ткани. Здесь хиломикроны взаимодействуют с липопротеинлипазой — ферментом, который освобождает жирные кислоты. Эти жирные кислоты поступают в жировую и мышечную ткани для накопления и окисления соответственно. После изъятия триглицеридного ядра остаточные хиломикроны содержат большое количество холестерина и апопротеина Е. Апопротеин Е специфически связывается со своим рецептором в клетках печени, после чего остаточный хиломикрон захватывается и катаболизируется в лизосомах. В результате этого процесса освобождается холестерин, который затем преобразуется в желчные кислоты и выводится или участвует в формировании новых липопротеинов, образующихся в печени (ЛПОНП). При нормальных условиях хиломикроны находятся в плазме в течение 1-5 ч. после приема пищи [2], [3].
Внутренний путь. Печень постоянно синтезирует триглицериды, утилизируя свободные жирные кислоты и углеводы. В составе липидного ядра ЛПОНП они выходят в кровь. Внутриклеточный процесс формирования этих частиц схож с таковым для хиломикронов, за исключением различия в апопротеинах. Последующее взаимодействие ЛПОНП с липопротеинлипазой в тканевых капиллярах приводит к формированию остаточных ЛПОНП, богатых холестерином (ЛППП). Примерно половина этих частиц выводится из кровотока клетками печени в течение 2-6 ч. Остальные претерпевают модификацию с замещением оставшихся триглицеридов эфирами холестерина и освобождением от всех апопротеинов, за исключением апопротеина В. В результате формируются ЛПНП, которые содержат ¾ всего плазменного холестерина. Их главная функция — доставка холестерина в клетки надпочечников, скелетных мышц, лимфоцитов, гонад и почек [3]. Модифицированные ЛПНП (окисленные продукты, количество которых возрастает при повышенном содержании в организме активных форм кислорода, так называемом окислительном стрессе) могут распознаваться иммунной системой как нежелательные элементы. Тогда макрофаги их захватывают и выводят из организма в виде ЛПВП. При чрезмерно высоком уровне ЛПНП макрофаги становятся перегруженными липидными частицами и оседают в стенках артерий, образуя атеросклеротические бляшки.
Основные транспортные функции липопротеинов приведены в таблице.
Таблица. Функции липопротеинов [5].
| Класс | Размеры | Функция |
|---|---|---|
| ЛПВП | 4-14 нм | Транспорт холестерина от периферийных тканей к печени |
| ЛПНП | 20-22,5 нм | Транспорт холестерина, триглицеридов и фосфолипидов от печени к периферийным тканям |
| ЛППП | 25-35 нм | Транспорт холестерина, триглицеридов и фосфолипидов от печени к периферийным тканям |
| ЛПОНП | 30-80 нм | Транспорт холестерина, триглицеридов и фосфолипидов от печени к периферийным тканям |
| Хиломикроны | 75-1200 нм | Транспорт холестерина и жирных кислот, поступающих с пищей, из кишечника в периферические ткани и печень |
Регуляция уровня холестерина
Уровень холестерина в крови в большой степени определяется диетой. Пищевые волокна снижают уровень холестерина, а пища животного происхождения повышает его содержание в крови.
Один из основных регуляторов метаболизма холестерина — рецептор LXR (рис. 3). LXR α и β принадлежат к семейству ядерных рецепторов, которые образуют гетеродимеры с ретиноидным Х рецептором и активируют гены-мишени. Их естественные лиганды — оксистерины (окисленные производные холестерина). Обе изоформы идентичны на 80% по аминокислотной последовательности. LXR-α обнаружен в печени, кишечнике, почках, селезенке, жировой ткани; LXR-β в небольшом количестве обнаруживается повсеместно [6]. Метаболический путь оксистеринов быстрее, чем у холестерина, и поэтому их концентрация лучше отражает краткосрочный баланс холестерина в организме. Существует всего три источника оксистеринов: ферментативные реакции, неферментативное окисление холестерина и поступление с пищей. Неферментативные источники оксистеринов как правило минорные, но в патологических состояниях их вклад возрастает (окислительный стресс, атеросклероз), и оксистерины могут действовать наряду с другими продуктами перекисного окисления липидов [6]. Основное влияние LXR на метаболизм холестерина: обратный захват и транспорт в печень, вывод с желчью, снижение кишечного всасывания. Уровень продукции LXR различается на протяжении аорты; в дуге, зоне турбулентности, LXR в 5 раз меньше, чем в участках со стабильным течением. В здоровых артериях повышение экспрессии LXR в зоне сильного потока оказывает антиатерогенное действие [7].
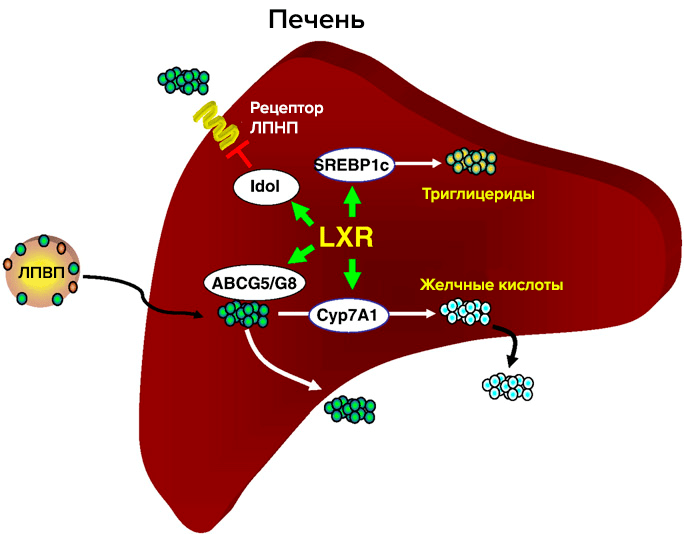
Рисунок 3. Участие рецептора LXR в метаболизме холестерина в печени
Важную роль в метаболизме холестерина и стероидов играет рецептор-«мусорщик» SR-BI (рис. 4). Он был обнаружен в 1996 году как рецептор для ЛПВП [8]. В печени SR-BI отвечает за избирательный захват холестерина из ЛПВП. В надпочечниках SR-BI опосредует избирательный захват этерифицированного холестерина из ЛПВП, который необходим для синтеза глюкокортикоидов. В макрофагах SR-BI связывает холестерин, что является первым этапом в обратном транспорте холестерина. SR-BI также захватывает холестерин из плазмы и опосредует его прямой выход в кишечник [9].
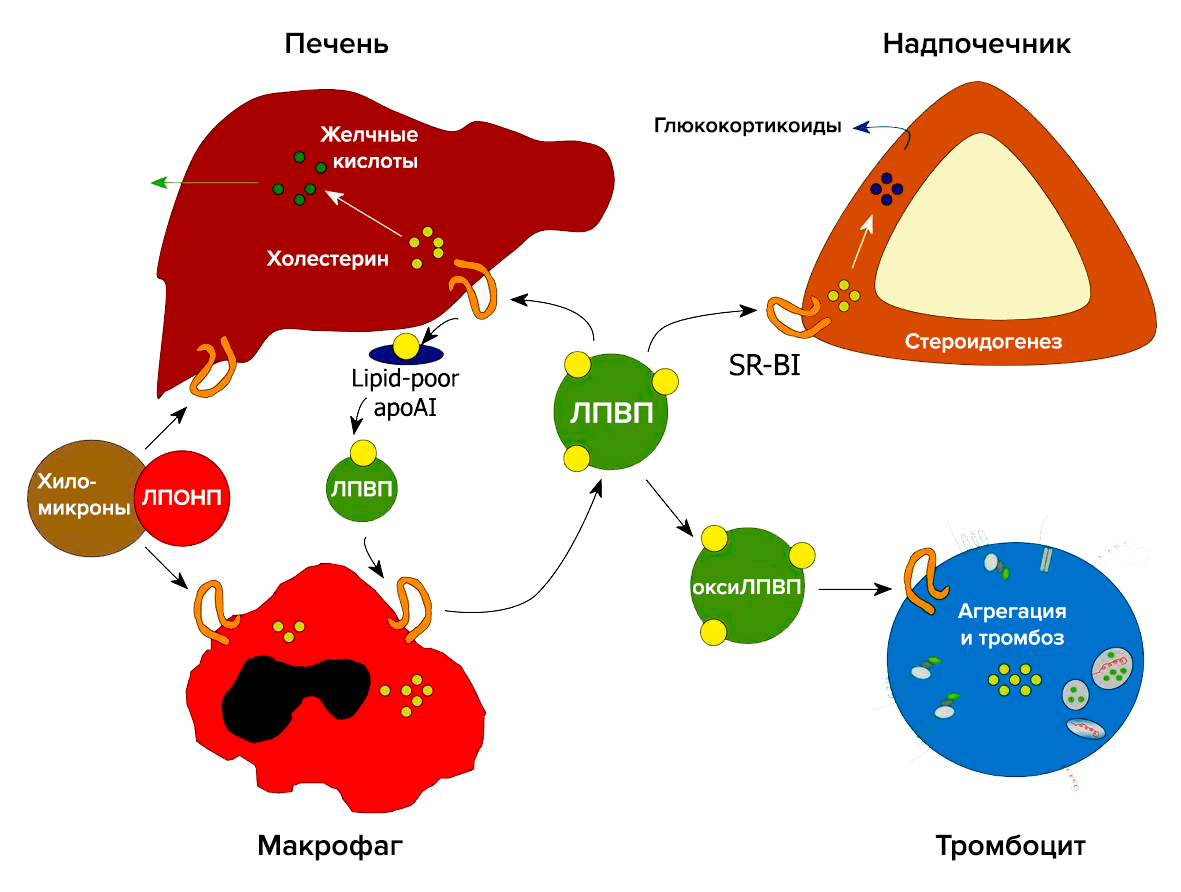
Рисунок 4. Участие рецептора SR-BI в метаболизме холестерина
Выведение холестерина из организма
Классический путь выведения холестерина: транспорт холестерина с периферии в печень (ЛПВП), захват клетками печени (SR-BI), экскреция в желчь и выведение через кишечник, где большая часть холестерина возвращается в кровь [10].
Основная функция ЛПВП — обратный транспорт холестерина в печень. Плазменные ЛПВП являются результатом комплекса различных метаболических событий. Состав ЛПВП очень различается по плотности, физико-химическим свойствам и биологической активности. Это сферические или дисковидные образования. Дисковидные ЛПВП в основном состоят из апопротеина A-I с вложенным слоем фосфолипидов и свободного холестерина. Сферические ЛПВП больше и дополнительно содержат гидрофобное ядро из эфиров холестерина и небольшого количества триглицеридов.
При метаболическом синдроме активируется обмен триглицеридов и эфиров холестерина между ЛПВП и триглицерид-богатыми липопротеинами. В результате содержание триглицеридов в ЛПВП повышается, а холестерина снижается (т.е. холестерин не выводится из организма) [11]. Отсутствие ЛПВП у людей встречается при болезни Tangier, главные клинические проявления которой — увеличенные оранжевые миндалины, роговичная дуга, инфильтрация костного мозга и мукозного слоя кишечника [3].
Если коротко обобщить, то страшен не сам холестерин, который является необходимым компонентом, обеспечивающим нормальную структуру клеточных мембран и транспорт липидов в крови, а кроме того он является сырьем для производства стероидных гормонов. Метаболические расстройства же проявляются при нарушении баланса ЛПНП и ЛПВП , что отражает нарушение системы транспорта липопротеинов, включающей работу печени, образование желчи и участие макрофагов. Поэтому любые заболевания печени, а также аутоиммунные процессы могут вызвать развитие атеросклероза даже при вегетарианской диете. Если мы вернемся к изначальным опытам Н.А. Аничкова по кормлению кроликов пищей, богатой холестерином, то увидим, что холестерин не встречается в естественном рационе кроликов и поэтому, как яд, нарушает работу печени, вызывает сильное воспаление сосудов и, как следствие, образование бляшек.
- Anitschkow N. and Chalatow S. (1983). Classics in arteriosclerosis re: On experimental cholesterin steatosis and its ificance in the origin of some pathological processes by N. Anitschkow and S. Chalatow, translated by Mary Z. Pelias, 1913. Arteriosclerosis, Thrombosis, and Vascular Biology. 3, 178-182;
- Климов А.Н. Причины и условия развития атеросклероза. Превентивная кардиология. М.: «Медицина», 1977. — 260-321 с.;
- Cox R.A. and Garcia-Palmieri M.R. Cholesterol, triglycerides, and associated lipoproteins. Clinical methods: the history, physical, and laboratory examinations (3rd Edition). Boston: Butter-worths, 1990. — 153-160 p.;
- Grundy S.M. (1978). Cholesterol bolism in man. West. J. Med. 128, 13-25;
- Википедия: «Липопротеины»;
- Wójcicka G., Jamroz-Wisniewska A., Horoszewicz K., Beltowski J. (2007). Liver X receptors (LXRs). Part I: Structure, , regulation of activity, and role in lipid bolism. Postepy Hig. Med. Dosw. 61, 736-759;
- Calkin A. and Tontonoz P. (2010). Liver X Receptor aling pathways and atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 30, 1513-1518;
- S. Acton, A. Rigotti, K. T. Landschulz, S. Xu, H. H. Hobbs, M. Krieger. (1996). Identification of Scavenger Receptor SR-BI as a High Density Lipoprotein Receptor. Science. 271, 518-520;
- Vrins C.L.J. (2010). From blood to gut: Direct secretion of cholesterol via transintestinal cholesterol efflux. World J. Gastroenterol. 16, 5953-5957;
- Van der Velde A.E. (2010). Reverse cholesterol transport: From classical view to new insights. World J. Gastroenterol. 16, 5908-5915;
- Wilfried Le Goff, Maryse Guerin, M.John Chapman. (2004). Pharmacological modulation of cholesteryl ester transfer protein, a new therapeutic target in atherogenic dyslipidemia. Pharmacology & Therapeutics. 101, 17-38;
- Липидный фундамент жизни;
- Наночастицами — по «плохому» холестерину!.
Источник