Синтез и регуляция гемоглобина
Содержание статьи
Синтез гемоглобина. Регуляция биосинтеза гемоглобина.
Мы поможем в написании ваших работ! Мы поможем в написании ваших работ! Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ? | Гем является небелковой частью многих гемопротеинов: · гемоглобин (до 85% общего количества гема организма), локализованный в эритроцитах и клетках костного мозга, · миоглобин скелетных мышц и миокарда (17%), · цитохромы дыхательной цепи, · ферменты цитохромоксидаза, цитохром P450, гомогентизатоксидаза, пероксидаза, миелопероксидаза, каталаза, тиреопероксидаза и т.д. — менее 1%. Синтез гема в основном идет в предшественниках эритроцитов, клетках печени, почек, слизистой кишечника, и в остальных тканях. Первая реакция синтеза с участием δ-аминолевулинат-синтазы (греч. δ — «дельта») происходит в митохондриях. Следующая реакция при участии аминолевулинатдегидратазы (порфобилиноген-синтазы) протекает в цитозоле. Здесь из двух молекул δ‑аминолевулиновой кислоты образуется циклический порфобилиноген (монопиррол). После синтеза порфобилиногена четыре его молекулы конденсируются с образованием уропорфириногена (тетрапиррол). Их различают два вида — уропорфириноген типа I и уропорфириноген типа III. В синтезе обоих видов порфиринов принимает участие уропорфириноген I-синтаза, в образования уропорфириногена III дополнительно принимает участие фермент уропорфириноген III-косинтаза. Судьба обоих типов уропорфириногена двояка: они могут окисляться до уропорфирина или декарбоксилироваться до копропорфириногена соответствующего типа. Копропорфириноген III превращается либо в копропорфирин III, либо идет на синтез протопорфирина IX. Копропорфириноген III окисляется в протопорфириноген IX и далее в протопорфирин IX. Последний после связывания с железом образует гем, реакцию катализирует феррохелатаза (гемсинтаза). Названия пигментов (уропорфирины и копропорфирины) были даны веществам по источнику их первоначального выделения, при этом восстановленные бесцветные формы называют порфириногенами. Для порфиринов характерно наличие изомерии вследствие различного расположения радикалов. Регуляция синтеза гема. 1. Скорость синтеза глобиновых цепей зависит от наличия гема, он ускоряет биосинтез «своих» белков. 2. Основным регуляторным ферментом синтеза гема является аминолевулинатсинтаза. · гем после взаимодействия с молекулой белка-репрессора формирует активный репрессорный комплекс, связывается с ДНК и подавляет транскрипцию, мРНК для фермента не образуется и синтез фермента прекращается. Также имеется отрицательный аллостерический эффект гема на фермент. · с другой стороны, достаточное количество ионов железа оказывает положительный эффект при синтезе молекулы аминолевулинатсинтазы. В клетке имеется особый железосвязывающий белок, который в отсутствии ионов железа обладает сродством к мРНК фермента и блокирует ее трансляцию в рибосоме, т.е. синтез белковой цепи. Ионы железа связываются с этим железосвязывающим белком, образуя с ним неактивный комплекс, что инициирует синтез фермента. 3. Положительным модулятором аминолевулинатсинтазы служит гипоксия тканей, которая в эритропоэтических тканях индуцирует синтез фермента. 4. В печени повышение активности аминолевулинатсинтазы вызывают соединения, усиливающие работу микросомальной системы окисления (жирорастворимые токсины, стероиды) — при этом возрастает потребление гема для образования цитохрома Р450, что снижает внутриклеточную концентрацию свободного гема. В результате происходит дерепрессия синтеза фермента. 70. Схема распада гемоглобина.Основная часть желчных пигментов образуется при распаде гема и гемоглобина в клетках ретикулоэндотелиальной системы (РЭС) и представляет собой многоступенчатый процесс: при окислении гемоглобина образуется вердоглобин; после отщепления глобина и железа образуется биливердин, который далее восстанавливается до билирубина. Билирубин — желто-красный пигмент, представляющий собой линейный тетрапиррол, токсичное, жирорастворимое вещество, способное нарушать окислительное фосфорилирование в клетках, в первую очередь в нервной ткани. В крови билирубин либо находится в свободном состоянии, либо в комплексе с альбумином (частично в виде альбумин-фосфатидного комплекса), в меньшем количестве — в комплексах с металлами, аминокислотами, пептидами и другими малыми молекулами. Образование таких комплексов предотвращает выделение билирубина с мочой. Такая форма билирубина называется свободной (неконъюгированной, несвязанной, непрямой). Она не дает прямой реакции с диазореактивом Эрлиха. Из сосудистого русла в гепатоциты билирубин попадает с помощью белка-переносчика (лигандина). В печени, при участии фермента УДФ‑глюкуронилтрансферазы, происходит реакция этерификации — взаимодействие OH‑группы глюкуроновой кислоты с карбоксильными группами билирубина и образование конъюгированного (связанного, прямого) билирубина, представляющего собой парное соединение с одним или двумя остатками глюкуроновой кислоты (моно- или диглюкуронид). Кроме глюкуроновой кислоты в реакцию могут вступать сульфаты, фосфаты, глюкозиды. В норме конъюгированный билирубин активно секретируется в желчные капилляры, где при участии β‑глюкуронидазы, вновь превращается в свободный билирубин и с током желчи попадает в тонкий кишечник. Здесь ферментами бактериальной флоры он восстанавливается до мезобилирубина и мезобилиногена (уробилиногена). Часть последних всасывается и с током крови вновь попадает в печень, где окисляется до ди‑ и трипирролов. При этом в здоровом организме в общий круг кровообращения и в мочу мезобилирубин и уробилиноген не попадают, а полностью задерживаются гепатоцитами. Невсосавшаяся часть пигментов ферментами бактериальной флоры толстого кишечника восстанавливается до стеркобилиногена и выделяется из организма, окрашивая кал. Незначительное количество стеркобилиногена через геморроидальные вены попадает в большой круг кровообращения, отсюда — в почки и выделяется с мочой. На воздухе стеркобилиноген и уробилиноген превращаются, соответственно, в стеркобилин и уробилин. |
Источник
94. Синтез гема и гемоглобина. Регуляция этих процессов. Вариации первичной структуры и свойств гемоглобина. Гемоглобинопатии.
Гем синтезируется во всех тканях, но с наибольшей скоростью в костном мозге и печени (рис. 13-2). В костном мозге гем необходим для синтеза гемоглобина в ретикулоцитах, в гепатоцитах — для образования цитохрома Р450.
Первая реакция синтеза гема — образование 5-аминолевулиновой кислоты из глицина и сук-цинил-КоА (рис. 13-3) идёт в матриксе митохондрий, где в ЦТК образуется один из субстратов этой реакции — сукцинил-КоА. Эту реакцию катализирует пиридоксальзависимый фермент аминолевулинатсинтаза.
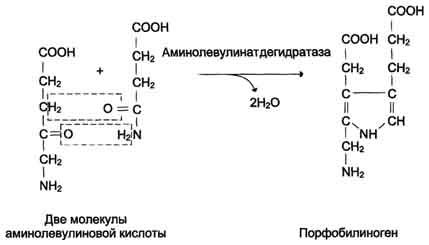
Из митохондрий 5-аминолевулиновая кислота поступает в цитоплазму. В цитоплазме проходят промежуточные этапы синтеза гема: соединение 2 молекул 5-аминолевулиновой кислоты молекулу порфобилиногена (рис. 13-4), дезаминированиепорфобилиногена с образованием гидроксиметилбилана, ферментативное превращение гидроксиметилбилана в молекулу уропор-фобилиногена III, декарбоксилирование последнего с образованием копропорфириногена III. Гидроксиметилбилан может также нефермента-тивно превращаться в уропорфириноген I, который декарбоксилируется в копропорфирино-ген I. Из цитоплазмы копропорфириноген III опять поступает в митохондрии, где проходят заключительные реакции синтеза гема. В результате двух последовательных окислительных реакций копропорфириноген III превращается в протопорфириноген IX, а протопорфириноген IX — в Протопорфирин IX. Фермент феррохела-таза, присоединяя к протопорфирину IX двухвалентноелентное железо, превращает его в гем (рис. 13-2). Источником железа для синтеза гема служит депонирующий железо белок ферритин. Синтезированный гем, соединяясь с α и β-полипепептидными цепями глобина, образует гемоглобин. Гем регулирует синтез глобина: при снижении скорости синтеза гема синтез глобина в ретикулоцитах тормозится.
Донором железа служит депонирующий железо в клетках белок ферритин.

Рис. 13-3. Реакция образования 5-аминолевулиновой кислоты.
Регуляция биосинтеза гема
Регуляторную реакцию синтеза гема катализирует пиридоксальзависимый фермент аминолевулинатсинтаза. Скорость реакции регулируется аллостерически и на уровне трансляции фермента.
Аллостерическим ингибитором и корепрессором синтеза аминолевулинатсинтазы является гем (рис. 13-5).
В ретикулоцитах синтез этого фермента на этапе трансляции регулирует железо. На участке инициации мРНК, кодирующей фермент, имеется последовательность нуклеотидов, образующая шпилечную петлю, которая называется железочувствительным элементом (от англ, iron-responsiveelement, IRE) (рис. 13-6).
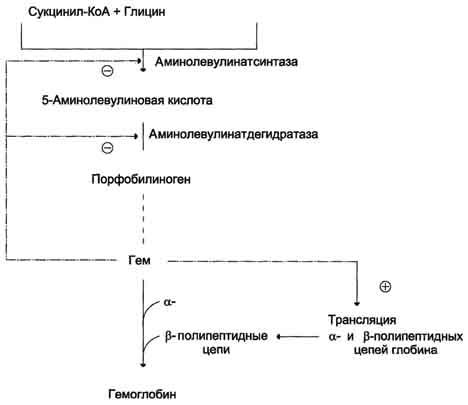
Рис. 13-5. Регуляция синтеза гема и гемоглобина. Гем по принципу отрицательной обратной связи ингибирует аминолевулинатсинтазу и аминолевулинатдегидратазу и является индуктором трансляции α- и β-цепей гемоглобина.
При высоких концентрациях железа в клетках оно образует комплекс с остатками цистеина регуляторного железосвязывающего белка. Взаимодействие железа с регуляторным железосвязывающим белком вызывает снижение сродства этого белка к IRE-элементу мРНК, кодирующей аминолевулинатсинтазу, и продолжение трансляции (рис. 13-6, А). При низких концентрациях железа железосвязывающий белок присоединяется к железо-чувствительному элементу, находящемуся на 5′-нетранслируемом конце мРНК, и трансляция аминолевулинатсинтазы тормозится (рис. 13-6, Б).
Аминолевулинатдегидратаза также аллостерически ингибируется гемом, но так как активность этого фермента почти в 80 раз превышает активность аминолевулинатсинтазы, то это не имеет большого физиологического значения.
Дефицит пиридоксальфосфата и лекарственные препараты, которые являются его структурными аналогами, снижают активность аминолевулинатсинтазы.
Альфа-полипептидная цепь заканчивается комбинацией аминокислот валина-лейцина, а бета-полипептидная цепь — комбинацией валина-гистидина-лейцина. Альфа- и бета-полипептидные цепи в гемоглобиновой молекуле не размещены линейно, как это выглядит на первый взгляд из данных,это первичная структура .
Гемоглобинопатии группа наследственных гемолитических анемий, обусловленных нарушениями первичной структуры гемоглобина (качественные гемоглобинопатии) или синтеза глобиновых цепей (количественные гемоглобинопатии).
Соседние файлы в папке экз
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник
Гем необходим для многих ферментов
Гем является небелковой частью многих гемопротеинов:
- гемоглобин (до 85% общего количества гема организма), локализованный в эритроцитах и клетках костного мозга,
- миоглобин скелетных мышц и миокарда (до 17%),
- цитохромы дыхательной цепи,
- ферменты цитохромоксидаза, цитохром P450, гомогентизатоксидаза, миелопероксидаза, каталаза и глутатионпероксидаза, тиреопероксидаза и т.д. — менее 1%.
Строение и синтез гема
Гем — структура, включающая в себя порфириновое кольцо (состоящее из 4 пиррольных колец) и иона Fe2+. Железо связывается с порфириновым кольцом двумя координационными и двумя ковалентными связями.
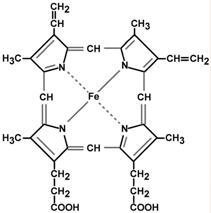
Строение гема
Синтез гема в основном идет в предшественниках эритроцитов, в клетках печени, почек, слизистой кишечника и в остальных тканях. Первая реакция синтеза с участием δ-аминолевулинат-синтазы (греч. δ — «дельта») происходит в митохондриях. Следующая реакция при участии аминолевулинатдегидратазы (порфобилиноген-синтазы) протекает в цитозоле, здесь из двух молекул δ‑аминолевулиновой кислоты образуется циклический порфобилиноген (монопиррол).
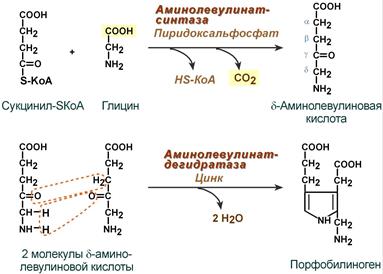
Синтез порфобилиногена
После синтеза порфобилиногена четыре его молекулы конденсируются в гидроксиметилбилан, который далее превращается в уропорфириноген типа I и уропорфириноген типа III. В синтезе обоих видов порфиринов принимает участие уропорфириноген I-синтаза, в образовании уропорфириногена III дополнительно принимает участие фермент уропорфириноген III-косинтаза.
Судьба обоих типов уропорфириногена двояка: они могут окисляться до уропорфирина (на рисунке не показано) или декарбоксилироваться до копропорфириногена соответствующего типа.
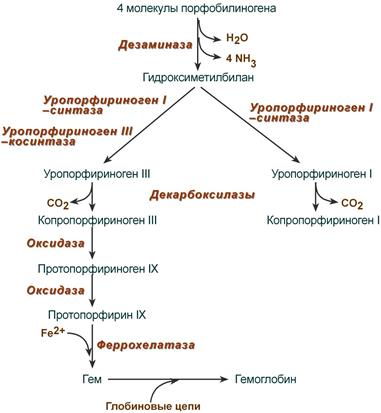
Синтез гема из порфобилиногена
Копропорфириноген III возвращается в митохондрии и окисляется в протопорфириноген IX и далее в протопорфирин IX. Последний после связывания с железом образует гем, реакцию катализирует феррохелатаза (гемсинтаза).
Скорость синтеза глобиновых цепей зависит от наличия гема, он ускоряет биосинтез «своих» белков.
Названия пигментов (уропорфирины и копропорфирины) были даны веществам по источнику их первоначального выделения, при этом восстановленные бесцветные формы называют порфириногенами. Для порфиринов характерно наличие изомерии вследствие различного расположения радикалов, что нашло отражение в порядковых номерах изомеров.
Регуляция синтеза гема
Основным регуляторным ферментом синтеза гема является аминолевулинатсинтаза.
1. Гем :
- напрямую оказывает отрицательный аллостерический эффект на фермент,
- влияет на транскрипцию фермента. После взаимодействия с молекулой белка-репрессора формирует активный репрессорный комплекс, связывается с ДНК и подавляет транскрипцию, мРНК для фермента не образуется и синтез фермента прекращается.
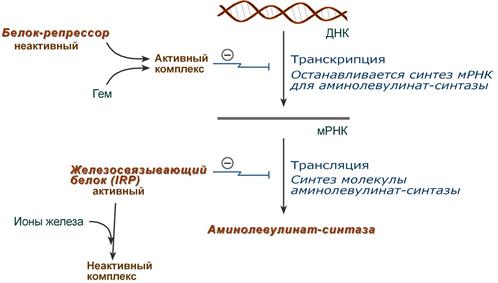
Регуляция синтеза аминолевулинатсинтазы
2. Ионы железа. Их достаточное количество оказывает положительный эффект при синтезе молекулы аминолевулинатсинтазы.
В клетке имеется железосвязывающий белок (англ. IRP, iron-responsive element-binding proteins — белок, связывающий железочувствительный элемент), который в отсутствии ионов железа обладает сродством к железочувствительному участку IRE (англ. iron-responsive element ) на матричной РНК фермента. Это связывание блокирует трансляцию мРНК в рибосоме, т.е. подавляет синтез белковой цепи.
При наличии ионов железа они связываются с железосвязывающим белком, образуя с ним неактивный комплекс, и это инициирует синтез фермента.
3. Положительным модулятором аминолевулинатсинтазы служит внутриклеточная гипоксия, которая в эритропоэтических тканях индуцирует синтез фермента.
4. В печени повышению активности аминолевулинатсинтазы способствуют различные соединения, усиливающие работу микросомальной системы окисления (жирорастворимые вещества, стероиды) — при этом возрастает потребление гема для образования цитохрома Р450, и снижается внутриклеточная концентрация свободного гема. В результате происходит усиление синтеза фермента.
Источник
Синтез гемоглобина
Синтезированный в митохондриях гем индуцируется синтез цепей глобина на полирибосомах. Гены цепей глобина расположены в 11 и 16 хромосоме.
Цепи глобина формируют глобулы и соединяются с гемом. 4 глобулы нековалентно соединяются в гемоглобин.
Гемоглобин начинает синтезироваться на стадии базофильного эритробласта, а заканчивается у ретикулоцитов. В ретикулоцитах также идет синтез пуринов, пиримидинов, фосфатидов, липида. Чувствительным биохимическим индикатором для отличия ретикулоцитов от зрелых клеток является утрата последними глутаминазы. Глутамин в ретикулоцитах — источник углерода для синтеза порфирина и азота для синтеза пурина.
Строение гемоглобина
Гемоглобин- тетрамерный хромопротеин, имеет массу 64,5кДа, состоит из 4 гемов и 4 глобинов. Глобины представлены полипептидными цепями различных типов,,,и т.д.-цепь содержит 141 АК, а- цепь — 146 АК. Отдельные участки полипептидных цепей образуют правозакрученные-спирали, особое расположение в пространстве которых формирует глобулы. Глобула -субъединицы содержит 8-спиралей, а-субъединицы -7. Гем располагается в щелях между Е иFспиралями глобина, прикрепляясь через гистидинF8 к спиралиFс помощью 5 координационной связи железа. Гидрофобные остатки аминокислот окружающие гем, препятствуют окислению железа водой. 4 глобулы с участием гидрофобных, ионных и водородных связей формируют шарообразный тетрамер гемоглобина. Максимально прочные связи, в основном за счет гидрофобных связей, образуются между- и-глобулами. В результате образуются 2 димера11и22. Димеры соединяются между собой в основном полярными (ионными и водородными) связями, поэтому взаимодействие димеров зависит от рН. Димеры легко перемещаются друг относительно друга. В центре тетрамера глобулы прилегают друг к другу неплотно, образуя полость.
Функции гемоглобина
Обеспечивают перенос кислорода от легких к тканям. В сутки около 600литров;
Участвует в переносе углекислого газа и протонов от тканей к легким;
Образует гемоглобиновый буфер, регулирует КОС крови.
Производные гемоглобина
Гемоглобин со свободной шестой координационной связью железа в составе гема называется апогемоглобином.
Шестая координационная связь может связывать различные лиганды, с образованием следующих производных гемоглобина:
оксигемоглобинHbО2(Fe2+) — соединение молекулярного кислорода с гемоглобином. Процесс называется оксигенацией; обратный процесс — дезоксигенацией.
карбоксигемоглобинHbСО (Fe2+). Связь гема с СО в двести раз прочнее, чем с О2. В норме в крови содержится 1%HbСО. У курильщиков к вечеру концентрацияHbСО достигает 20%. При отравлении СО, из-за недостаточного снабжения тканей кислородом может наступить смерть.
метгемоглобинHbОН (Fe3+). Образуется при воздействии на гемоглобин окислителей (оксидов азота, метиленового синего, хлоратов). В норме в крови содержится<1%HbОН. Накопление метгемоглобина при некоторых заболеваниях (например, нарушение синтеза ГЛ-6-фосфатДГ), отравлении окислителями может стать причиной смерти, так как метгемоглобин не способен к переносу кислорода;
цианметгемоглобинHbСN(Fe3+). Образуется при присоединении СN-к метгемоглобину. Эта реакция спасает организм от смертельного действия цианидов. Поэтому для лечения отравлений цианидами применяют метгемоглобинообразователи (нитритNa);
Карбгемоглобинобразуется, когда гемоглобин связывается с СО2. Однако СО2присоединяется не к гему, а кNН2- группам глобина, с образованием карбаматов:
HbNH2 +CO2=HbNHCOO-+H+
Карбгемоглобин выводит из организма 10-15% СО2.
ДезоксигемоглобинHb(Fe2+). Форма гемоглобина не связанная с кислородом. Дезоксигемоглобин связывает больше СО2, чем оксигемоглобин.
В цитохромахгем присоединяется к белковой части через 5 и 6 координационные связи железа (через гистидин и метионин Е иFспиралей). Занятость всех координационных связей не позволяет цитохромам присоединять лиганды, поэтому они могут переносить только по 1 электрону.
Механизм насыщения гемоглобина кислородом
Гемоглобин присоединяет О2последовательно, по одной молекуле на каждый гем.
В апогемоглобине,благодаря координационной связи с белковой частью, атом железа выступает из плоскости гема в направлении гистидинаF8.
Присоединение О2к шестой координационной связи железа вызывает его перемещение в плоскость гема, за ним перемещаются гистидинF8 и полипептидная цепь, в состав которой он входит.
Происходит изменение конформации текущего протомера и связанных с ним оставшихся протомеров. При этом у протомеров возрастает сродство к кислороду, в результате каждый следующий кислород присоединяется к гемоглобину лучше предыдущего. Четвертая молекула кислорода присоединяется к гемоглобину в 300 раз легче, чем первая молекула. Обратный процесс аналогичен, чем больше О2отдают протомеры, тем легче идет отщепление последующих молекул О2.
Кривая диссоциации кислорода для гемоглобина
Кооперативность в работе протомеров гемоглобина формирует сигмовидный характер кривой насыщения его кислородом в зависимости от парциального давления кислорода.
S-образная кривая насыщения гемоглобина кислородом имеет важное биологическое значение.
Во-первых, пологий участокS-образной кривой (выше 60мм.рт.ст.) обеспечивает максимальное насыщение гемоглобина кислородом в легких, даже если концентрация кислорода в альвеолярном воздухе заметно снижена. Например, в альвеолярной крови приРО2=95 мм.рт.ст. гемоглобин насыщается кислородом на 97%, а при РО2=60 мм.рт.ст. — на 90%.
Во-вторых,Крутой наклон среднего участка S-образной кривой (от 10 до40 мм.рт.ст.) обеспечивает максимальный переход кислорода от гемоглобина к тканям.
В области венозного конца капилляра приРО2 = 40 мм.рт.ст. гемоглобин насыщен кислородом на 73%. При снижении РО2 на 5 мм.рт.ст. насыщение гемоглобина кислородом уменьшается на 7%.
Аллостерическая регуляция насыщения гемоглобина кислородом
Кроме РО2 на насыщение гемоглобина кислородом влияют и другие факторы, например, рН, температура, давление, концентрация 2,3-ДФГ,РСО2.
Увеличение температуры, присоединение к гемоглобину Н+, 2,3-ДФГ, СО2уменьшает сродство гемоглобина к кислороду, при этом кривая диссоциации оксигемоглобина сдвигается вправо и гемоглобин легче отдает кислород тканям.
Эффект Бора
Влияние рН на характер кривой диссоциации оксигемоглобина называется эффектом Бора(по имени датского физиолога Христиана Бора, впервые открывшего этот эффект).
Гемоглобин в дезоксигенерированном состоянии имеет более высокое сродство к протонам, чем оксигемоглобин. Другими словами R — форма (оксигенерированная) является более сильной кислотой, чем Т-форма (дезоксигенерированная). Поэтому когда дезоксигемоглобин в легких присоединяет кислород, происходит переход в R — форму и разрыв некоторых связей, в результате чего и высвобождаются протоны, ответственные за эффект Бора. Наоборот, при высвобождении кислорода образуется Т-структура и разорванные связи между субъединицами должны быть восстановлены, и протоны вновь присоединяются к остаткам гистидина в - цепях. Таким образом, протонирование гемоглобина снижает его сродство к О2 и увеличивает потребление О2 в ткани.
Эффект Бора имеет важное физиологическое значение. Образующийся в тканях СО2 должен транспортироваться в легкие. Он поступает в эритроциты по градиенту напряжения. В них фермент карбоангидраза превращает его в Н2СО3, который диссоциирует на бикарбонат, ион и протон. Последний сдвигает равновесие влево в уравнении (1).
Hb + 4 O2= Hb (О2)4 + (H+)n
Где n — величина порядка 2; число зависит от целого комплекса параметров, тем самым заставляя Hb О2 отдавать свой кислород.
НСО3- пассивно продвигается через ионный канал по градиенту концентрации в сыворотку.
Продвижение НСО3- не сопровождается перемещением Н+, поскольку нет канала, позволяющего ему пройти через мембрану эритроцитов. Для сохранения ионного равновесия при выходе НСО3- из клетки, Cl- перемещаются внутрь её через тот же ионный канал. Такое двойное перемещение известно как хлоридный сдвиг (сдвиг Хамбургера).
Растворенный НСО3- движется вместе с венозной кровью обратно в легкие. Здесь высвобождение протона из гемоглобина при оксигениции приводит к образованию НСО3- (по принципу Ле-Шателье).
НСО3-+ Н+= Н2СО3-,
что позволяет карбоангидразе образовать СО2.
Разрушение НСО3- в эритроците обуславливает вхождение в него НСО3- из сыворотки, так что в легких происходит обратный хлоридный сдвиг, приводящий к выведению СО2 с выдыхаемым воздухом.
Аллостерическая регуляция сродства гемоглобина к кислороду 2,3-ДФГ
2,3-ДФГ снижает сродство гемоглобина к кислороду и, таким образом, повышает отдачу кислорода тканям. Если кровь израсходовала весь свой запас ДФГ, гемоглобин остается фактически насыщенным кислородом. При акклиматизации в условиях высокогорья содержание ДФГ в эритроцитах резко увеличивается. ДФГ является аллотерическим лигандом, так как связывается с гемоглобином в другом по сравнению с О2 участком. ДФГ встраивается в полость тетрамерной молекулы гемоглобина, полость образована остатками всех 4 протомеров.
В Т — форме (дезоксигенерированной) молекулы Hb имеются дополнительные связи, и поэтому размер центральной полости больше, чем в R — форме (дезоксигемоглобине). Поэтому ДФГ взаимодействует только с Т — формой стабилизируя её, путем образования связи между атомами кислорода ДФГ и тремя положительно заряженными группами в каждой из - цепей.
В легких при высоком парциальном давлении кислород взаимодействует с Hb, изменяется конформация белка, уменьшается центральная полость и ДФГ вытесняется из гемоглобина.
Виды гемоглобинов
Гемоглобины различаются по белковой части. Бывают физиологические и аномальные виды гемоглобинов. Физиологические образуются на разных этапах нормального развития организма, а аномальные — вследствие нарушения последовательности аминокислот в глобине физиологических видов гемоглобина.
Физиологические виды гемоглобина
1) эмбриональные гемоглобины(Gover I, Gover II). На ранних этапах развития плода в первые недели развития, когда в желточном мешке возникают очаги кроветворения начинается синтез-цепей (эпсилон). Из четырёхцепей образуется гемоглобин Gover I. Затем у эмбриона, длина которого не превышает 2,5см, начинается синтез-цепей, которые вместе-цепями образуют гемоглобин Gover II (22). Затем синтез-цепей прекращается и Gover гемоглобины полностью исчезают у трехмесячного эмбриона. Если они остаются у новорожденного, то это признак врожденной аномалии развития.
2) фетальный гемоглобин- HbF (от латинского fetus — плод). Фетальный гемоглобин сменяет эмбриональные гемоглобины, вместо эпсилон — цепей (- цепей) начинают синтезироваться гамма-цепи (- цепи). HbF состоит из 2и 2цепей. HbF — является главным гемоглобином плода и составляет к моменту рождения 50-80% всего гемоглобина. HbF имеет более высокое сродство к кислороду, что позволяет ему забирать кислород от гемоглобина матери и передавать его тканям плода. Эта особенность связана с низким сродством HbF к 2,3-ФГК.
Кроме перечисленных основных видов гемоглобинов плода, у здорового плода выделяются и другие виды гемоглобинов: например, гемоглобин Bart`s, (4), Portland-1 (S22).
Схема электрофореза
гемоглобина здорового плода
+
А1
F
А2
Gower I
Bart`s
Portland -1
Gower II
3) гемоглобин А1- тетрамер (22) составляет около 98% гемоглобина эритроцитов взрослого человека. Начинает синтезироваться на 8 месяце развития плода.
4) гемоглобин А2- тетрамер (22). Его содержание в эритроцитах взрослого человека равно 2%. Гемоглобин А2, также как и гемоглобин F, обладает более высоким сродством к кислороду по сравнению с гемоглобином А1.
5) гемоглобин А3(22) образуется по мере старения эритроцита, при присоединении к цистеину-цепи глутатиона.
6) гемоглобин А1С- гликозилированный гемоглобин А.
Аномальные виды гемоглобинов
Аномальные гемоглобины возникают в результате мутации генов, кодирующих ицепи. Известно несколько сотен мутантных гемоглобинов человека (в большинстве случаев функционально активных).
Таблица №1 замена аминокислот в ипептидных цепях гемоглобина
тип гемоглобина | нормальный остаток и его положение в цепи | замена |
С | глу 6 в - цепи | лиз |
Д | лей 28 в - цепи | глу |
Е | глу 26 в - цепи | лиз |
G | глу 43 в - цепи | ала |
GpH | асл 68 в - цепи | лиз |
J | лиз 16 в - цепи | асл |
М | вал 67 в - цепи | глу |
О | глу 116 в - цепи | лиз |
S | глу 6 в - цепи | вал |
Болезни гемоглобинов
Болезни гемоглобинов называют гемоглобинозами, их насчитывают более 200.
Гемоглобинозы делятся на гемоглобинопатии и таласемии.
Гемоглобинопатии, возникают в результате точечных мутаций в структурных генах, кодирующих полипептидные цепи гемоглобина. Поэтому в крови появляется аномальный гемоглобин.
Серповидноклеточная анемия- классический пример наследственной гемоглобинопатии. В норме в-субъединицах гемоглобина в шестом положении находится гидрофильная глутаминовая кислота. В гемоглобине S глутаминовая кислота заменена на гидрофобный валин. Такая замена приводит к появлению на поверхности-субъединицы гидрофобного («липкого») участка, который соединяется с гидрофобным карманом другой молекулы гемоглобина S. Происходит полимеризация гемоглобина S и его осаждение в виде длинных волокон. Длинная волокнистая структура нарушает нормальную форму эритроцитов, превращая её из двояковогнутого диска в серповидную, которая имеет тенденцию блокировать капилляры. Такие эритроциты преждевременно разрушаются, способствуя развитию анемии. Если поражены обе гомологичные хромосомы, заболевание может оказаться смертельным. Заболевание широко распространено в географических зонах, где наиболее часто встречается злокачественная форма малярии. Высокий показатель заболеваемости можно объяснить положительной селекцией генома носителей аномальных генов. Серповидная красная кровяная клетка «неудобна» для развития малярийного плазмодия.
Существенное ухудшение состояния больных наблюдается в условиях высокогорья при низких давлениях кислорода. Это связано с тем, что полимеризоваться способна только дезоксиформа S гемоглобина. Так как в молекуле оксиформы S-гемоглобина нет гидрофобного кармана («липкого участка»), и она не способна к полимеризации.
Талассемия- генетическое заболевание, обусловленное отсутствием или снижением синтеза одной из цепей гемоглобина. При данном заболевании отсутствуют дефекты в структурных генах, кодирующих,,,-цепи.
Причиной талассемий являются мутации генов-операторов, контролирующих транскрипцию структурных генов ,,,-цепей гемоглобина.
В результате несбалансированного образования глобиновых цепей образуются тетрамеры гемоглобина, состоящие из одинаковых протомеров.
В зависимости от того, формирование какой глобиновой цепи нарушается, выделяют , , , — талассемии.
Талассемии делятся так же на гомозиготныеигетерозиготные.
Гомозиготная -талассемия- формирование-цепи полностью подавляется. Симптомы заболевания появляются приблизительно через полгода после рождения, когда происходит полное переключение синтеза-цепи гемоглобина F на-цепь. У ребенка прогрессирует анемия. Увеличиваются селезенка и печень. Лицо приобретает монголоидные черты (из-за чрезмерного разрастания костного мозга скулы выдаются вперед, нос приплюснут), при рентгенологическом исследовании черепа наблюдается феномен «игл ежа» («hair — standing -on -end»). В попытке восполнить эритроциты, утраченные в результате не эффективного эритропоэза и увеличении гемолиза, ткани черепа, чрезмерно разрастаясь и гипертрофируясь, порождают такое изменение медуллярной пластинки.
α-талассемия — недостаток образования α-глобиновых цепей приводит к нарушению образования HbF у плода. Избыточные γ-цепи образуют тетрамеры, называемые гемоглобином Барта. Этот гемоглобин при физиологических условиях имеет повышенное сродство к кислороду и не проявляет кооперативных взаимодействий между протомерами. В результате гемоглобин Барта не обеспечивает развивающийся плод необходимым количеством кислорода, что приводит к тяжёлой гипоксии. При α-талассемии отмечают высокий процент внутриутробной гибели плода. Выжившие новорождённые при переключении с γ- на β-ген синтезируют β-тетрамеры или НbН, который, подобно гемоглобину Барта, имеет слишком высокое сродство к кислороду, менее стабилен, чем НbА и быстро разрушается. Это ведёт к развитию у больных тканевой гипоксии и к смерти вскоре после рождения.
Для всех этих заболеваний характерны некоторые общие закономерности:
1). нарушаются пропорции в составе гемоглобина крови. Например, при - талассемии в крови появляется 15% гемоглобина А2, 15 — 60% гемоглобина F;
2). эритроциты приобретают не нормальную форму (мишеневидную, каплевидную). Такие эритроциты в пределах 1 дня захватываются ретикулярной соединительной тканью (например, селезенкой) и подвергаются распаду (по этой причине селезёнка оказывается гипертрофированной), что приводит к развитию гемолитической анемии.
Соседние файлы в папке 7 модуль
- #
- #
- #
- #
- #
- #
- #
Источник