Синтез липидов и холестерина в клетке
Содержание статьи
интез фосфолипидов в клетке — цик
Синтез фосфолипидов в клетке — цикл Кеннеди• Основные клеточные фосфолипиды синтезируются в мембране ЭПР, главным образом, со стороны цитозоля. • Регуляция синтеза новых липидов контролируется локализацией ферментов, участвующих в этом процессе. • Биосинтез холестерина регулируется протеолизом фактора транскрипции, который интегрирован в мембрану ЭПР. Наряду с транслокацией и подготовкой белков к секреторному процессу, эндоплазматический ретикулум (ЭПР) функционирует как место первичного образования клеточных фосфолипидов. Эти молекулы синтезируются в мембране эндоплазматического ретикулума (ЭПР) и затем распределяются по отдельным мембранам и органеллам внутри клетки. К их числу относится плазматическая мембрана, митохондрии и органеллы секреторного пути. При возникновении необходимости клетка должна обладать способностью увеличивать количество мембран. Наиболее очевидно это проявляется при удвоении плазматической мембраны и всех органелл, которое происходит в ходе клеточного цикла. Наряду с этим, при гиперфункции, отдельные органеллы могут увеличиваться в размерах. Например, при созревании предшественников В-лимфоцитов в плазматические клетки, секретирующие антитела, ЭПР сильно увеличивается, что соответствует потребностям усиленного транспорта белков по секреторному пути. Образование фосфолипидов из растворимых предшественников de novo происходит, главным образом, на мембране ЭПР со стороны цитоплазмы, в ходе процесса, называемого цикл Кеннеди. Как показано на при росте мембраны ЭПР две молекулы жирных кислот, связанные с ацетил КоА, реагируют с глицерин-3-фосфатом, образуя диацилглицерин (ДАТ). В отличие от своих предшественников, ДАГ достаточно гидрофобен для того, чтобы интегрироваться в мембрану ЭПР. Попав в мембрану, ДАГ взаимодействует с «головой» будущей молекулы фосфолипида. Как показано на рисунке ниже, основные фосфолипиды различаются, главным образом, природой «головы» молекулы. Все они образуются при присоединении «головы» к ДАГ. Вначале происходит ее фосфорилирование, а затем присоединение к цитидиндифосфату (ЦДФ). После этого «голова» с одной из фосфатных групп переносится на ДАГ, и синтез завершается. Присоединение головной группы холина к ЦЦФ с образованием фосфатидилхолина иллюстрирует использование клеткой субклеточной локализации белков для регуляции процессов биосинтеза. В этой реакции участвует фермент цитидилтрансфераза (ЦТ), и она контролирует скорость всего процесса синтеза фосфатидилхолина. В клетке ЦТ находится в двух различных формах: в неактивной, в цитозоле, и в активной, с цитозольной стороны мембраны ЭПР. Механизм, который обеспечивает существование двух этих форм фермента, представляется не вполне ясным. Очевидно, однако, что условия внутри клетки, которые сигнализируют о необходимости усиления синтеза фосфатидилхолина, могут контролировать переход ЦТ из одной формы в другую. Например, увеличение в клетке концентрации свободного ДАТ или жирных кислот вызывает миграцию ЦТ на мембрану ЭПР, где эти компоненты реагируют с холином. В то же время, когда в клетке содержится много фосфатидилхолина, ЦТ локализуется в цитозоле. Таким образом, предотвращается синтез фосфатидилхолина. Наряду с фосфатидилхолином, который является наиболее распространенным мембранным фосфолипидом, в цикле Кеннеди также образуются фосфатидилэтаноламин и фосфатидилинозитол. У дрожжей S. cerevisiae синтезируется фосфатидилсерин. У животных, однако, этот фосфолипид образуется за счет другой реакции, которая не требует участия ЦЦФ, однако также протекает с цитозольной стороны мембраны ЭПР. Интересная особенность синтеза фосфатидилэтаноламина заключается в том, что хотя этот фосфолипид образуется в цикле Кеннеди, он также может синтезироваться в митохондриях. В этих органеллах он образуется при модификации фосфатидилсерина, который образовался в ЭПР. Образование фосфатидилэтаноламина в клеточных компартментах, отличающихся от тех, в которых синтезируется его предшественник, предполагает существование механизма транспорта липидов между двумя компартментами. Имеются данные, позволяющие предполагать, что этот транспорт осуществляется посредством специального субдомена ЭПР, который называется мембраной, связанной с митохондрией (МСМ). МСМ представляет собой участок ЭПР, который физически ассоциирован с митохондрией. Он характеризуется плоской поверхностью и тесным контактом с органеллой. В МСМ находятся ферменты синтеза фосфатидилсерина. Возможно, что область контакта мембраны с митохондрией обеспечивает быстрый перенос фосфатидилсерина в органеллу, хотя механизм этого переноса остается неясным. Наряду с участием в синтезе фосфолипидов, эндоплазматический ретикулум (ЭПР) является местом синтеза стеринов. К числу основных стеринов мембран клетки относится холестерин; его биосинтез осуществляется в несколько этапов. Первые несколько реакций происходят в цитозоле, а остальные в мембране ЭПР. Также в ЭПР находятся элементы, необходимые для регуляции образования холестерина. Ключевыми медиаторами этого регуляторного пути являются белки, связывающиеся с регуляторным стеринреспонсивным элементом (SREBP). Эти белки обычно интегрированы в мембрану эндоплазматического ретикулума (ЭПР) и со стороны люмена содержат небольшую петлю, соединяющую два трансмембранных домена, N- и С-концевые участки которых ориентированы в сторону цитозоля. Также в ЭПР находится белок SCAP (белок, активирующий разрезание SREBP). Этот белок контролирует уровень внутриклеточного холестерина. Когда возникает необходимость в повышенном синтезе холестерина, SCAP транспортирует SREBP из ЭПР в аппарат Гольджи. Там происходит разрезание SREBP, и N-терминальный участок молекулы высвобождается в цитозоль. (В реакции участвуют те же протеазы, которые расщепляют АТФ 6 при возникновении стрессорных условий в ЭПР. Затем N-терминальный домен SREBP мигрирует в ядро, где действует как фактор транскрипции, активируя экспрессию генов в каскаде реакций биосинтеза холестерина.
Новая растворимая в липидах молекула включается в мембрану. Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021 — Также рекомендуем «Механизм транспорта липидов между органеллами в клетке» Оглавление темы «Синтез белка в клетке»:
|
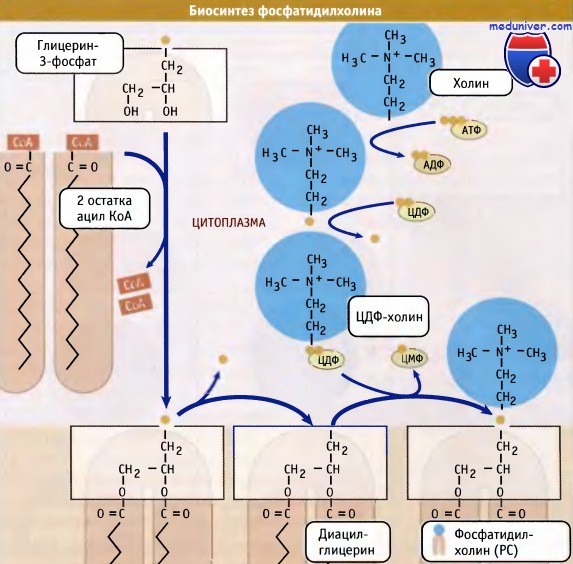 При объединении водорастворимых цитоплазматических компонентов (глицерин-3-фосфата, ацил КоА и полярной головки) образуется молекула фосфолипида.
При объединении водорастворимых цитоплазматических компонентов (глицерин-3-фосфата, ацил КоА и полярной головки) образуется молекула фосфолипида.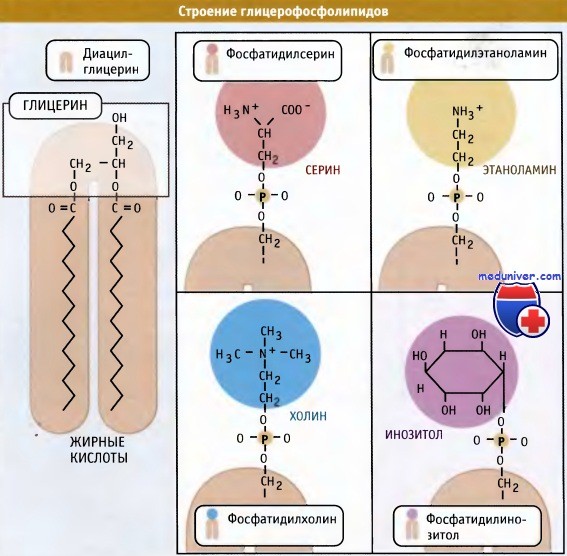 Основные мембранные фосфолипиды различаются по составу головок.
Основные мембранные фосфолипиды различаются по составу головок.Источник
Биосинтез холестерина
Биосинтез холестерина — образование в живом организме органического спирта холестерина стероидной природы. Синтез холестерина происходит в клетках печени (50 %), кишечнике и коже. В клетке он идёт в гладком эндоплазматическом ретикулуме и цитозоле. Биосинтез холестерина служит основой синтеза других стероидных соединений. Начальные этапы синтеза являются общими с этапами синтеза других изопреноидов.
Нумерация атомов в молекуле холестерина
В 1940-е годы Блох с сотр. установил, что все атомы углерода холестерина происходят из ацетата, причём оба атома включаются в одинаковых количествах. В настоящее время установлена следующая цепь биосинтеза холестерина, включающая в себя несколько ступеней. (Образование промежуточных соединений этого пути может происходить и другими способами).
- Превращение трёх молекул активного ацетата в пятиуглеродный мевалонат.
- Превращение мевалоната в активный изопреноид — изопентенилпирофосфат.
- Образование тридцатиуглеродного изопреноида сквалена из шести молекул изопентенилдифосфата.
- Циклизация сквалена в ланостерин.
- Последующее превращение ланостерина в холестерин.
Синтез мевалоната[править | править код]
Синтез мевалоната протекает в три этапа.
- Образование ацетоацетил-КоА из двух молекул ацетил-КоА с помощью тиолазного фермента ацетоацетилтрансферазы. Реакция обратима. Происходит в цитозоле.
- Образование β-гидрокси-β-метилглутарил-КоА из ацетоацетил-коА с третьей молекулой ацетил-КоА с помощью гидроксиметилглутарил-КоА-синтазы (ГМГ-КоА-синтазы). Реакция также обратима. Происходит в цитозоле.
- Образование мевалоната восстановлением ГМГ и отщеплением HS-KoA с помощью НАДФ-зависимой гидроксиметилглутарил-КоА-редуктазы (ГМГ-КоА-редуктаза). Происходит в гЭПР. Это первая практически необратимая реакция в цепи биосинтеза холестерина, а также она лимитирует скорость биосинтеза холестерина. Отмечены суточные колебания синтеза этого фермента. Активность его увеличивается при введении инсулина и тиреоидных гормонов, снижается при голодании, введении глюкагона, глюкокортикоидов.
Схема этапа:
Существуют и другие пути синтеза мевалоната.
Синтез изопентенилпирофосфата[править | править код]
1, 2. Вначале мевалоновая кислота дважды фосфорилируется с помощью АТФ: до 5-фосфомевалоната, а затем до 5-пирофосфомевалоната. 3. 5-пирофосфомевалонат фосфорилируется по 3 атому углерода, образуя нестабильный промежуточный продукт — 3-фосфо-5-пирофосфомевалонат. 4. Последний декарбоксилируется и дефосфорилируется, образуется изопентенилпирофосфат.
Синтез сквалена[править | править код]
- Изопентенилпирофосфат изомеризуется в диметилаллилпирофосфат.
- Конденсация изопентенилпирофосфата (С5) с диметилаллилпирофосфатом (С5) и образование геранилпирофосфата (С10). При этом высвобождается молекула пирофосфата.
- Конденсация изопентенилпирофосфата (С5) с геранилпирофосфатом (С10). Образуется фарнезилпирофосфат (С15) и высвобождается ещё одна молекула пирофосфата.
- Конденсация двух молекул фарнезилпирофосфата (С15) «голова-к-голове» и образование сквалена (С30). Реакция проходит с затратой НАДФН, и высвобождаются две молекулы пирофосфата. Молекулы фарнезилпирофосфата конденсируются концами, несущими пирофосфатные группы. Сначала отщепляется одна пирофосфатная группа и образуется промежуточный прескваленпирофосфат. Он, в свою очередь, восстанавливается с помощью НАДФН. Второй пирофосфат уходит. Образуется сквален.
Начиная со сквалена, продукты пути биосинтеза холестерина нерастворимы в водной среде и участвуют в дальнейших реакциях, будучи связанными со стеринпереносящими белками (СПБ).
Синтез ланостерина[править | править код]
1. Под действием скваленэпоксидазы образуется эпоксид сквалена. 2. Затем эпоксид сквалена циклизуется в ланостерин. При этом метильная группа у С14 переносится на С13, а метильная группа у С8 — на С14.
Синтез холестерина[править | править код]
Ланостерин превращается в мембранах гладкого эндоплазматического ретикулума в холестерин.
- Метильная группа при С14 окисляется, и образуется 14-десметилланостерин.
2. Затем удаляются ещё два метила при С4, и образуется зимостерол. 3. Далее двойная связь C8=С9 перемещается в положение С8=С7 и образуется Δ7,24-холестадиенол. 4. Двойная связь далее перемещается в положение С5=С6,образуется десмостерол. 5. После чего в боковой цепи восстанавливается двойная связь, и образуется холестерин.
(Восстановление двойной связи в боковой цепи может, однако, происходить и на предшествующих стадиях биосинтеза холестерола.)
Другие пути синтеза[править | править код]
У некоторых организмов при синтезе стероидов могут встречаться другие варианты реакций (например, немевалонатный путь образования пятиуглеродных молекул).
Литература[править | править код]
- [www.xumuk.ru/biologhim/169.html Химическая энциклопедия сайта www.humuk.ru. Скан учебника «Биологическая химия», Берёзов Т. Т., Коровкин Б. Ф.]
- Кольман Я., Рём К.-Г., «Наглядная биохимия», пер. с нем., М., «Мир», 2009.
- Марри Р., Греннер Д., Мейес П., Родуэлл В., «Биохимия человека». М., «Мир», 1993.
Промежуточные продукты метаболизма холестерина и стероидов | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Мевалонатный путь |
| ||||||||||||||
| Не-мевалонатный путь |
| ||||||||||||||
| В холестерин |
| ||||||||||||||
| Витамин D С-27: Холестаны |
| ||||||||||||||
| Жёлчные кислоты С-24: Холаны |
| ||||||||||||||
| Стероидные гормоны |
| ||||||||||||||
| Не у человека |
| ||||||||||||||
Примечания Н — нейростероидные гормоны см. также ферменты, заболевания | |||||||||||||||
Некоторые внешние ссылки в этой статье ведут на сайты, занесённые в спам-лист. Эти сайты могут нарушать авторские права, быть признаны неавторитетными источниками или по другим причинам быть запрещены в Википедии. Редакторам следует заменить такие ссылки ссылками на соответствующие правилам сайты или библиографическими ссылками на печатные источники либо удалить их (возможно, вместе с подтверждаемым ими содержимым). Список проблемных ссылок
|
Источник
Вопрос 1. Классификации органелл.
7
Тема: «Основы цитофизиологии.»
2. Органеллы- постоянные клеточные структуры, расположенные в гиалоплазме. Их подразделяют на органеллы общего значения (обязательно есть во всех клетках) и специальные органеллы (реснички, жгутики, микроворсинки, миофибриллы, акросома), которые встречаются лишь в некоторых типах клеток.
По строению выделяют мембранные и немембранные органеллы. Существует 5 мембранных органелл общего значения (митохондрии, эндоплазматическая сеть — ЭПС, комплекс Гольджи — кГ, лизосомы и пероксисомы) и 2 немембранных — рибосомы и клеточный центр.
В зависимости от функций, органеллы принято распределять по функциональным системам (аппаратам) клетки.
Вопрос 2. Синтетический аппарат клетки: рибосомы, эндоплазматическая сеть, комплекс Гольджи.
Синтетический аппарат клеткиобеспечивает синтезы различных веществ и включает ЭПС, кГ и рибосомы.
Рибосомы- округлые мелкие, немембранные органеллы, состоящие из двух округлых субъединиц — малой и большой. Каждая субъединица образованы рибосомальной РНК и сложным набором белков. Синтез рРНК и сборка субъединиц происходит в ядрышке, а их объединение — уже в цитоплазме. Рибосомы обеспечивают процесс трансляции белка. Малая субъединица связывается с иРНК, а большая катализирует образование пептидных связей между аминокислотами.
Одиночные рибосомы неактивны и для белкового синтеза они объединяются в цепочки, нанизываясь на молекулу иРНК. Так образуются полисомы.Свободные полисомы синтезируют белки, которые диффузно распределяются в гиалоплазме.
Синтез мембранных белков, лизосомальных белков и секреторных белков, которые будут выведены за пределы клетки, осуществляют полисомы, прикрепленные к ЭПС.
При синтезе секреторных и лизосомальныз белков используется особый механизм, который позволяет полипептидной цепи проникать внутрь полости ЭПС. После завершения синтеза молекула приобретает вторичную и третичную структуру и выйти наружу уже не может. Синтез таких белков начинается с особого участка — сигнального пептида. Благодаря ему рибосома соединяется с рибофорином — белком, который встроен в мембрану ЭПС. В присоединении участвует еще и специальный мембранный рецептор ЭПС. После присоединения рибосомы рибофорин приобретает форму канала, через который проходит синтезируемая полипептидная цепь. Когда белковая молекула готова, сигнальный участок отсоединяется.
Если синтезируется мембранный белок, то в полипептидной цепи оказывается еще один участок, который заякоривает белковую цепь в мембране.
ЭПС представляет собой сложную систему мембранных полостей. Обычно в форме плоских цистерн, распределенных по всей клетке.
Есть два типа ЭПС — гранулярная и агранулярная. К поверхности грЭПС прикрепляются полисомы.
Итак, главные функции грЭПС: синтез, химическая модификация, накопление и транспортировка белков.
Агранулярная ЭПСявляется продолжением грЭПС, но лишеная ЭПСявляется продолжением грЭПС, но лишена рибосом и имеет иной набор белков-ферментов. аЭПС — трубчатыми каналами. У нее множество функций:
Синтез липидов и холестерина, поэтому ее много в клетках, синтезирующих стероидные гормоны и жиры.
Синтез гликогена (клетки печени)
Детоксикация вредных веществ (лекарственные препараты, алкоголь, токсины)
Накопление Са2+, необходимого для сокращения мышечных клеток.
От ЭПС отшнуровываются транспортные пузырьки, содержащие синтезированные вещества, перемещаются в сторону комплекса Гольджи и сливаются с ним.
Комплекс Гольджи- мембранная органелла, представленная диктиосомами (стопка из 3-10 плоских цистерн). Диктиосома имеет незрелую поверхность, обращенную к ЭПС (цис-) и зрелую, обращенную к плазмолемме (транс-). С цис-поверхностью сливаются транспортные мембранные пузырьки, содержащие продукты синтеза, которые отшнуровываются от ЭПС. Вещества, попавшие в полости кГ, направляются в различные части диктиосомы, где подвергаются процессингу. Это химические превращения молекул — к ним могут присоединяются сахара, сульфатные и фосфатные группы, белковые молекулы могут частично расщепляться и т.д.
От боковых участков кГ отшнуровываются гидролазные пузырьки, заполненные гидролитическими ферментами. Из них формируются лизосомы.
Функции кГ:
Синтез полисахаридов и гликопротеинов (слизь, гликокаликс).
Процессинг молекул
Накопление продуктов синтеза, их упаковка и транспортировка.
Формирование лизосом.
Вопрос 3.Аппарат внутриклеточного перевариваниявключает эндосомы и лизосомы и обеспечивает расщепление крупных молекул.Эндосомы- это мембранные пузырьки, которые образуются при эндоцитозе (фаго- или пиноцитозе). Они содержат протеазы, которые в условиях слабокислой среды осуществляют мягкое, ограниченное расщепление захваченных продуктов. Прежде всего, здесь распадаются крупные комплексы: гормон-рецептор, антиген-антитело и т.д., а при дальнейшем закислении до рН — 5.5 начинается более глубокое переваривание. Эндосомы должны слиться с гидролазными пузырьками. Гидролазные пузырьки (первичные лизосомы) — мелкие мембранные вакуоли, которые отшнуровываются от кГ и содержат различные гидролитические ферменты: протеазы, нуклеазы, липазы в неактивной форме.
После этого слияния образуется лизосома (эндолизоса, вторичная лизосома). Благодаря мембранному ферменту Н+АТФазе, которая работает как протонная помпа, внутри лизосомы среда быстро становится кислой. Ее ферменты переходят в активную форму и расщепляют макромолекулы до низкомолекулярных веществ, которые через мембрану диффундируют в гиалоплазму.
Различают несколько видов лизосом:
Фаголизосомаосуществляет гетерофагию, т.е. переваривает материал, захваченный при фагоцитозе. Гетерофагия — важнейший механизм, который снабжает клетку питательными веществами и используется клетками защитной системы организма — макрофагами и нетрофилами.
Аутофаголизосомапереваривает собственные компоненты клетки, подлежащие замене. Поэтому происходит постоянное обновление клеточных структур. Например, в нейронах 70-летнего человека большинство органелл не старше 1 месяца.
Остаточное тельце- содержит непереваренный материал, который так и хранится внутри клетки или выделяется за ее пределы. Пример — липофусциновые гранулы, содержащие «пигмент старения», которые постепенно накапливаются в долгоживущих клетках — нейронах, кардиомиоцитах и мешают их работе.
Соседние файлы в папке Лекции (общ)
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник