Сродство гемоглобина к оксиду углерода больше чем к кислороду в
Содержание статьи
Влияние двуокиси углерода и водородных ионов на сродство гемоглобина к кислороду и кислородную емкость крови.
Обеспечение кислородного режима организма при широком диапазоне колебаний р02 и рС02 в воде и различном функциональном состоянии рыб происходит в результате согласованной работы систем дыхания и кровообращения, а также изменения количества и функциональных свойств гемоглобина — дыхательного белка, способного обратимо связывать кислород в органах дыхания и отдавать его в активнодышащих тканях. Иными словами, снабжение организма кислородом на необходимом уровне при изменении его содержания в воде или кислородного запроса обеспечивается сложной физиолого-биохимической системой, в которой объединены органы дыхания, кровообращения и гемоглобин, определяющий кислородную емкость крови. Хотя кислород способен физически растворяться в плазме крови, но емкость этого «носителя» (0,5 мл/100 мл) значительно уступает кислородной емкости гемоглобина рыб (5 — 10 мл/100 мл крови). При этом следует иметь в виду, что кислородная емкость крови рыб, как правило, пропорциональна содержанию в ней гемоглобина и потому существенно колеблется от вида к виду (от 2,4 — 4,5 до 15,7 — 19,7 мл/100 мл крови). Однако кислородная емкость крови определяется не только количеством гемоглобина, но и рядом других факторов, влияющих на степень его сродства к гемоглобину, прежде всего содержанием двуокиси углерода и величиной рН крови.
Основным показателем сродства гемоглобина к кислороду служит концентрация кислорода в воде, а точнее, его напряжение (р02), при котором половина молекул гемоглобина насыщена кислородом (Я50 — напряжение полунасыщения) и находится в форме оксигемоглобина (НЬ02), а другая остается ненасыщенной (НЬ). Зависимость количества оксигемоглобина, а точнее, соотношение между гемоглобином и оксиге-моглобином при каждой данной концентрации кислорода в воде графически выражается кривыми кислородной диссоциации, которые еще называют кривыми кислородного насыщения. Как правило, эти кривые имеют сигмоидную форму, что отражает кооперативное взаимодействие субъединиц гемоглобина при связывании кислорода, и описываются уравнением
где у — процент гемоглобина, связанного с кислородом.
Чем ниже содержание кислорода в воде, при котором происходит полунасыщение (Psо), тем выше сродство гемоглобина к кислороду и наоборот. При высоком сродстве гемоглобина к кислороду кривые диссоциации оксигемоглобина сдвинуты влево, а при более низком — вправо. Гемоглобин с пониженным сродством к кислороду отдает его в тканях легче, чем гемоглобин с повышенным сродством к кислороду. Помимо р02, в воде многие другие факторы оказывают существенное влияние на связывание кислорода гемоглобином и кривые его диссоциации. Особое место среди них принадлежит С02 и другим кислотам, оказывающим влияние на рН крови.
Поступление двуокиси углерода в кровяное русло ведет к подкислению крови (снижению величины рН) и снижению сродства гемоглобина к кислороду.
Кривая кислородной диссоциации смещается вправо, т. е. при повышенной концентрации С02 имеет место повышенная отдача кислорода при любом заданном его напряжении в активнодышащих тканях.
Это явление называется прямым эффектом Бора. Оно имеет ярко выраженное адаптивное значение: при поступлении С02 из тканей в капилляры гемоглобин отдает больше кислорода, чем это было бы без двуокиси углерода. Эффект Бора облегчает доставку тканям дополнительного кислорода, потребность в котором увеличивается при различных функциональных нагрузках на организм. Вместе с тем в жабрах рыб по мере отдачи С02 связывание кислорода гемоглобином увеличивается. Показателем выраженности эффекта Бора служит изменение величины Pso на единицу рН.
Наряду с прямым эффектом Бора существует обратный эффект, или эффект Холдена, т. е. повышение сродства гемоглобина к С02 при его дезоксигенации в тканях и облегчение отдачи С02 крови при ее оксигенации в жабрах. Оба этих эффекта связаны с кислотными свойствами гемоглобина.
Мы уже отмечали, что оксигемоглобин — более сильная кислота, чем дезоксигемоглобин, поскольку при оксигенации кислые группы белка каждой из субъединиц гемоглобина снижают рК. При соединении гемоглобина с кислородом его отрицательный заряд увеличивается, вытесняется соответствующее количество ионов НСОз и облегчается отдача С02.
Повышенное содержание С02 в крови снижает не только сродство гемоглобина к кислороду, но и общую кислородную емкость крови — эффект Рута. При подкислении крови избытком С02 или других кислот снижается величина максимального насыщения крови кислородом при любом р02. Эффект Рута, вызванный избытком С02, наступает быстрее, чем вызванный эквивалентным количеством молочной кислоты.
Таким образом, избыток С02, ведущий к снижению рН крови, оказывает существенное влияние на функциональные свойства гемоглобина и прежде всего на степень его сродства к кислороду и к самой двуокиси углерода. Интересно, что гиперкапния оказывает на сродство гемоглобина к кислороду и кислородную емкость крови рыб более существенное воздействие, чем у млекопитающих. Между тем системы дыхания и кровообращения у наземных позвоночных и, в частности у млекопитающих, более чувствительны к избытку С02, чем у рыб. Из этого следует, что в адаптивных перестройках физиолого-биохимической системы обеспечения кислородного режима организма рыб важнейшая роль принадлежит биохимической компоненте этой системы, т. е. изменению функциональных свойств гемоглобина. Видимо, поэтому влияние двуокиси углерода и рН крови на кривые диссоциации гемоглобина и кислородную емкость крови у многих видов рыб выражены сильнее, чем у млекопитающих. Правда, выраженность эффектов Бора и Рута у разных по экологии видов рыб существенно различается. У рыб, обитающих в малопроточных, обедненных кислородом водах, гемоглобин которых обладает высоким сродством к кислороду, двуокись углерода оказывает менее выраженное воздействие на кривые диссоциации, изменения которых лежат в физиологических пределах.
Так, например, у карпа, сома, ильной рыбы при рС02, равном 10 мм рт.ст., напряжение полунасыщения гемоглобина составляет около 10 мм рт.ст., в то время как у высокоподвижных и оксифильных рыб (3 вида форели, чукучан) аналогичное повышение напряжения двуокиси углерода в воде приводит к увеличению Р50 гемоглобина до 35 мм рт.ст. Даже при значительно меньшем увеличении напряжения С02 (от 2 до 10 мм рт.ст.) у рыб с высоким Рso кривые диссоциации оксигемоглобина могут быть резко сдвинуты вправо. Углекислота, облегчая отдачу кислорода тканям, значительно затрудняет насыщение гемоглобина кислородом в жабрах. В результате у рыб может развиться гипоксия даже при нормальном содержании кислорода в воде. У высокоактивных рыб, таких, как форель, при мышечной работе в крови появляется больше молочной кислоты, чем у малоактивных (сом, карп), поэтому эффект Бора у первых выражен сильнее, чем у вторых. Эти молекулярные особенности гемоглобина благоприятны для функционирования в экстремальных условиях, когда активнодышащие ткани нуждаются в повышенном притоке кислорода и возникает необходимость высвобождать его. Вместе с тем повышенная чувствительность гемоглобина к ионам Н+ у активных рыб имеет и свою вторую, но уже негативную сторону, поскольку при длительном интенсивном плавании, т. е. при напряженной работе, в кровяное русло поступает огромное количество молочной кислоты. В этих случаях, при снижении рН крови, эффект Бора будет угнетать процесс связывания гемоглобином кислорода в жабрах, что может привести и, подчас, приводит даже к гибели рыб от асфиксии. Поэтому чрезвычайный интерес представляют пока еще малочисленные данные о наличии у некоторых видов рыб двух типов гемоглобина: чувствительного к ионам Н+ с ярко выраженным эффектом Бора и нечувствительного к ионам Н+, не проявляющего эффект Бора. Гемоглобин первого типа в присутствии высоких концентраций С02 теряет способность связывать кислород, а второго сохраняет ее. У японского угря один из двух выявленных типов гемоглобина — Ei обладает повышенным сродством к кислороду {Р$о — 2,1 мм рт.ст.) и не обнаруживает эффекта Бора, а второй тип гемоглобина — ?2 имеет пониженное сродство к кислороду (Р%о — 14 мм рт.ст.) и выраженный эффект Бора. Количественно преобладает второй тип гемоглобина (Ei : Е2 = 3 : 7).
Мы разделяем предположение, что гемоглобин без эффекта Бора служит для системы транспорта 02 важным аварийным резервом, который может быть использован в любом случае, когда основные формы гемоглобина будут испытывать сильную отрицательную модуляцию в результате метаболического ацидоза.
Снижение общей кислородной емкости крови под влиянием С02 и подкисление крови иного происхождения (эффект Рута) проявляется у многих рыб, но не у всех. Выраженный эффект Рута отмечен, как правило, у высокоактивных рыб, таких, как форель, скумбрия, морской петух. Так, например, увеличение рС02 до 10 мм рт.ст. снижает кислородную емкость крови у морского петуха на 40%. При рС02, равном 30 мм рт.ст., насыщение крови кислородом у морского петуха снижается почти на 60%, у опсануса на 45%, у скумбрии на 30%. У некоторых малоподвижных рыб (скат, рыба-прилипала) эффект Рута отсутствует. У двоякодышащей рыбы Neoceratodus эффект Бора есть, а эффект Рута отсутствует, у акулы Squalus отсутствуют оба эффекта. Все эти данные свидетельствуют о том, что выраженность влияния избытка двуокиси углерода, так же как и концентрации водородных ионов, на сродство гемоглобина к кислороду и кислородную емкость крови неодинаковы у разных видов рыб и во многом определяются особенностями их экологии.
Физиолого-биохимические изменения при низких рН. Концентрация водородных ионов в воде является важнейшим экологическим эффектом, изменение которого оказывает существенное влияние на функциональное состояние и биохимический статус рыб. Повышение концентрации Н+ (снижение величины рН) в пресноводных водоемах вследствие усиленного поступления С02 органического происхождения (в ночные часы), сброса кислых стоков в водоем или выпадения «кислых дождей» ведет к развитию разных по направленности, длительности, выраженности и обратимости полифункциональных физиолого-биохимических изменений в организме рыб.
Еще в 30-х годах было установлено, что снижение величины рН и «закисление»воды вызывают у многих видов пресноводных рыб (девя-тииглая колюшка, карась, гольян, форель, карп и 5 видов окуней) угнетение дыхания, снижение интенсивности потребления кислорода и повышение его пороговой концентрации. Так, например, при рН, равном 7,0, гипоксия у голубого окуня развивается при концентрации кислорода 2,25 мл/л, а снижение величины рН всего лишь на единицу (до 6,0) — уже при 5 мл/л 02.
По данным Т. В. Новиковой, интенсивность потребления кислорода у карпа снижается с 0,24 до 0,16, а у окуня с 0,12 до 0,10 мг 02 на 1 г живой массы за 1 ч. Из этого следует, что окунь значительно более устойчив к изменениям величины рН, чем карп.
Как мы уже отмечали в предыдущих разделах, снижение величины рН, вызванное избыточным содержанием двуокиси углерода или лактата в крови, а также под влиянием любых других причин приводит к снижению сродства гемоглобина к кислороду. При этом кривые диссоциации смещаются вправо и изменяется их сигмоидная форма. Снижение сродства гемоглобина к кислороду и кислородной емкости крови (эффекты Бора и Рута) неизбежно ведет к развитию эндогенной гипоксии даже при нормальном содержании кислорода в воде. Свидетельством тому могут служить экспериментальные данные об увеличении содержания гемоглобина, количества эритроцитов и молочной кислоты в крови рыб при кислотном стрессе.
Так, например, под влиянием низких рН (3,0-4,0) у форели происходит увеличение концентрации гемоглобина на 29%, количества эритроцитов на 23% и гематокрита на 17%. Однонаправленные изменения концентрации гемоглобина и числа эритроцитов, способствующие увеличению кислородной емкости крови, вполне обоснованно рассматриваются в качестве компенсаторной реакции на возникающую у рыб под влиянием низких рН гипоксию.
При снижении рН воды до 4,2 у ручьевой форели отмечено повышение напряжения двуокиси углерода в артериальной крови, величины гематокрита и увеличение вентиляции жабр. Существует, однако, мнение, что в отсутствие С02 гипервентиляция жабр у лососевых рыб выражена слабо или не проявляется вообще.
X. Эллис был, по-видимому, первым, кто отметил повреждающее действие низких рН на жабры, а также усиленное образование, накопление и коагуляцию жаберной слизи.
Образующаяся при этом на жаберном эпителии коагуляционная пленка служит дополнительным препятствием для нормального газообмена вследствие увеличения диффузионного барьера. В дальнейшем гистопатологические изменения жаберной ткани рыб (гиперемия, деструкция и отслоение клеток жаберного эпителия от эндотелия капилляров) и усиленную секрецию слизи при остром и подостром кислотном отравлении отмечал каждый, кто изучал этот вопрос.
Нам известна всего лишь одна работа, в которой авторам лочему-то не удалось обнаружить патологических изменений жаберной ткани и образование слизи у ручьевой форели даже к моменту гибели после 7,5 часового воздействия остролетальной величины рН, равной 3,4. Это тем боле странно, что у такого высокоустойчивого ко многим стрессорным воздействиям вида рыб, каким является серебряный карась, уже в течение первого часа при рН 4,0 отмечено выраженное разрушение жаберного эпителия. В проверочной тщательно выполненной работе на том же виде рыб — на ручьевой форели было показано, что уже при рН 5,2 происходит гипертрофия слизистых клеток жаберного эпителия, а на жабрах накапливается слизь. В дальнейшем с увеличением кислотности воды до 3,5 отмечено разрушение жаберного эпителия и его отторжение от опорных клеток. Накопление слизи на жабрах в период, когда дыхание особенно затруднено, отмечено и у других видов лососевых рыб.
Объективный анализ накопленных экспериментальных данных позволяет считать твердо установленным, что первичной точкой приложения действия низких величин рН или «органом-мишенью» являются жабры и их ведущая функция — газообмен, т. е. обеспечение организма кислородом и освобождение от избытка двуокиси углерода.
Однако обеспечение дыхательного гомеостаза является важнейшей, но не единственной функцией жабр.
Наряду с почками жабры играют активную роль в поддержании водно-солевого гомеостаза, обеспечивая ионное и осмотическое постоянство внутренней среды. Обитая в резко различающихся по концентрации неорганических ионов морских и пресных водах, рыбы вынуждены с помощью различных морфофизиологических и физиолого-биохимических механизмов постоянно решать проблему стабилизации химического состава внутренней среды, т. е. изолировать внутриклеточные и внеклеточные жидкости от прямого влияния повышенной, или пониженной, солености окружающей среды. Основой решения этой проблемы служит непроницаемость для воды и ионов поверхность тела рыб. Исключение составляют именно жабры, непроницаемость которых сделала бы невозможной обеспечение рыб кислородом. Между тем через жабры идет постоянная «утечка» ионов из организма пресноводных рыб. По современным данным экстраренальные потери достигают 75-90% солей.
Для компенсации неизбежных потерь ионов рыбы располагают механизмом активного переноса ионов против концентрационного градиента из окружающей среды в организм или из организма в окружающую среду. Эти ионные насосы (главным образом натриевый) работают за счет энергии гидролиза АТФ. В энергетическом отношении это чрезвычайно дорогое физиолого-биохимическое приспособление (до 30% всех энергозатрат), однако оно обеспечило саму возможность существования рыб в условиях резких ионных градиентов между организмом и средой. Локализованы ионные насосы в уязвимых точках поверхности тела, т. е. в жабрах, где происходит интенсивный трансэпителиальный ионный обмен между организмом и средой. Особо следует подчеркнуть, что транспорт растворенных веществ через эпителий всегда сопряжен с переносом воды, в связи с чем проблема поддержания водно-солевого равновесия решается совместной деятельностью двух основных осморегуляторных органов — жабр и почек.
Также рекомендуем прочитать:
- Двуокись углерода и концентрация водородных ионов — буферная емкость крови
- Влияние избытка двуокиси углерода на систему обеспечения кислородного режима организма рыб
- Неясен и механизм ингибирующего влияния высоких концентраций водородных ионов на активный транспорт натрия
- Степень сродства гемоглобина к кислороду у рыб
- В условиях научно-технического прогресса города оказывают все большее влияние
- Влияние гстероауксииа на рост корней
- Влияние дождевых червей на структуру и химический состав почв
- Влияние животного населения на растительные сообщества глубоко осветили А. Г. Воронов и М. С
- Влияние изменений температуры на вегетативные процессы у морских рыб
- Влияние измененного гидробиологического режима водоема на жизнедеятельность рыб
- Влияние измененного гидрохимического режима водоема на жизнедеятельность рыб
- Влияние светового голодания на жителей крайнего Севера и Заполярья
- Влияние средовых условий на проявление комбинационной способности образцов томата
- Влияние средовых условий на эффективность отбора и экологическую стабильность генотипов томата
- Влияние среды на эффективность отбора в ранних поколениях
Источник
Гемоглобин: роль в газообмене и процессе дыхания
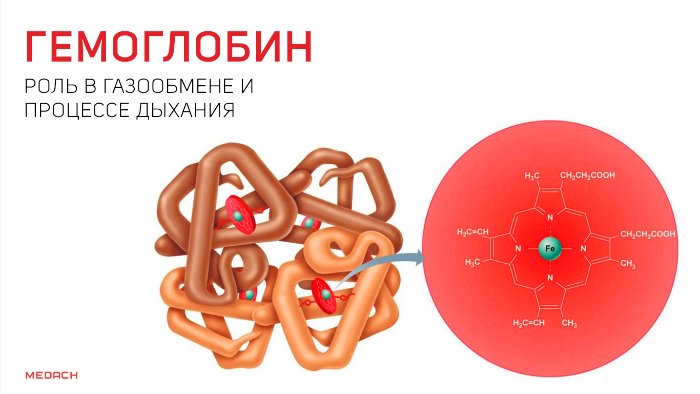
Одним из самых сложных процессов, что происходят в организме человека, несомненно, является дыхание. И сложность эта не только в танце легких, благодаря которому человек получает кислород, но и в процессах, с помощью которых этот кислород проникает дальше, в ткани, где превращается в углекислый газ, что отправляется в обратное путешествие. О данных процессах и пойдет речь далее.
Итак, приступим. Человек делает вдох, иии… Далеко не весь кислород поступает в легкие, а затем и в кровь. Часть вдыхаемого воздуха остается в так называемом мертвом пространстве. Мертвое пространство, в свою очередь, делится на анатомическое (дыхательные пути), в котором остается около 30 % вдыхаемого воздуха, и функциональное (вентилируемые, но по каким-то причинам не перфузируемые альвеолы).
Ухудшение альвеолярного газообмена может происходить при неглубоком и частом дыхании (причиной может стать перелом ребер, паралич дыхательной мускулатуры различного генеза и др.), а также при увеличении мертвого пространства, вызванном разнообразными причинами (нарушение перфузии альвеол в результате воспалительных заболеваний легких, удаление доли или целого легкого и др.), при снижении скорости кровотока по альвеолярным капиллярам (ТЭЛА, инфаркт легкого), при наличии диффузионного барьера (отек легких) и в результате ослабления альвеолярной вентиляции при обтурации просвета бронха. Газообмен между легкими и кровью происходит путем диффузии в соответствии с законом Фика. В легочных капиллярах она происходит за счет разности парциальных давлений в альвеолах и эритроцитах.
В альвеолах парциальное давление кислорода значительно превышает таковое для углекислого газа и составляет примерно 13,3 кПа (100 мм рт. ст.) и 5,3 кПа (40 мм рт. ст.) соответственно. Альвеолы омываются приносимой легочными артериями венозной кровью, в которой соотношение парциальных давлений этих двух газов обратно пропорционально и составляет приблизительно 5,3 кПа (40 мм рт. ст.) для кислорода и 6,1 кПа (46 мм рт. ст.) для углекислого газа. В среднем разница парциальных давлений составляет около 8 кПа (60 мм рт. ст.) для кислорода и около 0,8 кПа для углекислого газа.
Как уже было сказано выше, кислород путем диффузии проникает в кровь легочных капилляров. Диффузионное расстояние для кислорода при этом составляет 1–2 мкм, то есть именно на такое расстояние он проникает внутрь капилляра. Обмен крови в легочном капилляре происходит примерно за 0,75 секунды, но этого времени хватает на то, чтобы парциальные давления в альвеолах и в крови пришли в равновесие.
Кровь, в которой показатели парциального давления для кислорода и углекислого газа примерно равны таковым в альвеолах, называется артериализированной. Однако за счет наличия в легких артериовенозных шунтов и притока венозной крови из бронхиальных вен такой она остается недолго. В результате парциальное давление кислорода в аорте составляет примерно 12,0 кПа (как уже было сказано выше, парциальное давление в артериализированной крови равно таковому в альвеолах и составляет 13,3 кПа), а давление углекислого газа меняется незначительно и не приводит к затруднению его диффузии из крови в альвеолы.
Но кислород непосредственно в ткани попадает лишь в крайне незначительных количествах: для свободного перемещения по организму ему необходим транспортер. Эту функцию выполняет содержащийся в эритроцитах белок — гемоглобин. Гемоглобин существует в оксигенированной и неоксигенированной формах. В дезокси-гемоглобине железо находится на уровне порфиринового кольца и стабилизируется электростатическими силами, что обеспечивает поддержание всей структуры. Появившись, кислород начинает «тянуть» за железо, которое переносится на проксимальный гистидин на другом конце полипептидной цепи и меняет структуру всего протеина.
В результате гемоглобин переходит в оксигенированную форму, альфа- и бета-цепи при этом поворачиваются относительно друг друга на 15 градусов, облегчая присоединение остальных молекул кислорода. В итоге каждый из четырех содержащихся в нем атомов двухвалентного железа обратимо связывается с молекулой кислорода, что превращает молекулу гемоглобина в оксигемоглобин. По сравнению с миоглобином гемоглобин имеет низкое сродство к кислороду, однако оно не статично. Так, миоглобин может связывать кислород только одним участком, поэтому кривая его связывания — гипербола. Кривая связывания гемоглобина с кислородом имеет S-образную форму, демонстрируя, что при его связывании с первой молекулой кислорода гемоглобин имеет очень низкое сродство к кислороду, но при связывании последующих молекул кислорода сродство остальных его субъединиц к нему значительно увеличивается и в конечном счете повышается примерно в 500 раз.
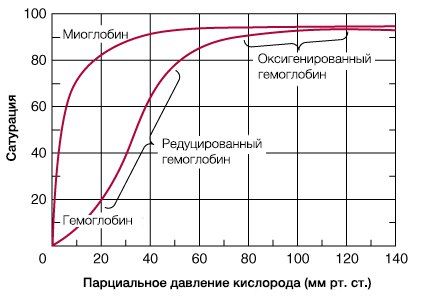
При этом альфа-цепи связывают кислород легче, чем бета-цепи. Этот процесс назван кооперативным взаимодействием. По мере снижения парциального давления кислорода в крови происходит его высвобождение из гемоглобина и поступление в ткани. Например, парциальное давление кислорода в работающих мышцах составляет всего 26 мм рт. ст, и при прохождении эритроцитов через капилляры, кровоснабжающие мышцы, происходит высвобождение и поступление в мышечные клетки примерно ⅓ всего переносимого гемоглобином кислорода. При повышении температуры тела также возрастает потребность в кислороде, что, в свою очередь, стимулирует высвобождение и поступление его в ткани. При снижении температуры, напротив, развивается гипоксия тканей, способствующая компенсаторному увеличению сродства гемоглобина к кислороду.
Гемоглобин также осуществляет перенос от тканей к легким продуктов тканевого дыхания — углекислого газа и ионов водорода. В ходе окислительных процессов в клетке выделяется углекислый газ, в результате гидратации которого образуются ионы водорода, что, в свою очередь, приводит к снижению рН. Давно известно, что снижение рН и повышение концентрации углекислого газа в крови оказывает сильное влияние на способность гемоглобина связывать кислород.
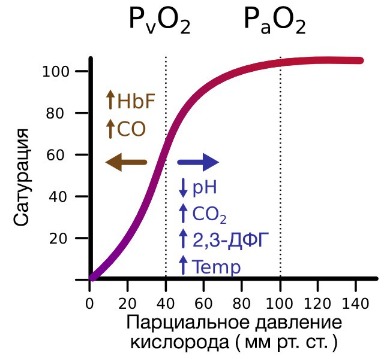
В периферических сосудах показатели рН низкие, и по мере связывания гемоглобина с ионами водорода и углекислым газом происходит снижение его сродства к кислороду. Это влияние величины рН и концентрации углекислого газа на способность гемоглобина связывать кислород называют эффектом Бора.
Обратная ситуация имеет место в альвеолярных капиллярах, где присоединение кислорода к гемоглобину превращает тот в более сильную кислоту.
При этом сродство гемоглобина к углекислому газу снижается, а повышение кислотности гемоглобина приводит к высвобождению излишка ионов водорода и образованию в крови из бикарбоната угольной кислоты, которая затем распадается на воду и углекислый газ. В обоих случаях углекислый газ из крови поступает в альвеолы, а затем в атмосферу. Данный процесс назван эффектом Холдейна. Стоит отметить, что важную роль в образовании углекислого газа в эритроцитах играет ион хлора, поступающий в плазму крови в обмен на бикарбонат при участии белка-переносчика АЕ1. Данный процесс в англоязычной литературе получил название «Chloride shift» или «перенос Хамбургера».
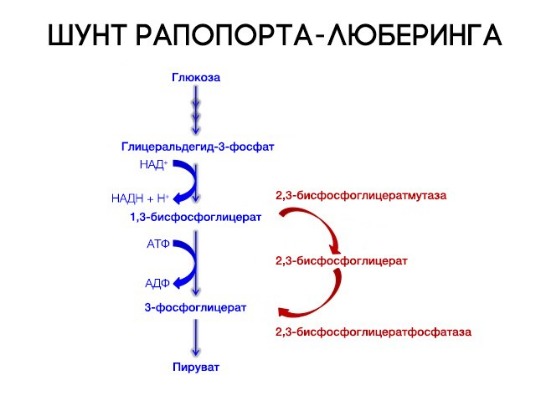
На сродство гемоглобина к кислороду оказывает влияние и присутствующее в эритроцитах вещество, получившее название 2,3-бисфосфоглицерат (БФГ). Его образование — своего рода побочная реакция анаэробного гликолиза, происходящего в эритроцитах в ходе ферментативного превращения глюкозы в пируват под действием фермента бифосфоглицератмутазы. БФГ способен самостоятельно связываться с неоксигенированной формой гемоглобина, образуя солевой мостик между двумя его бета-субъединицами и снижая сродство к кислороду.
При этом гемоглобин способен связать только одну молекулу БФГ, а при присоединении кислорода БФГ вытесняется из полости. В обычных условиях в эритроцитах крови содержится достаточно большое количество БФГ, которое может увеличиваться в условиях гипоксии (например, у дайверов при погружении на глубину), а также при восхождении на большую высоту. В первые часы подъема концентрация БФГ в эритроцитах будет возрастать, а сродство кислороду снижаться. Но на большой высоте парциальное давление будет значительно ниже такового на уровне моря, а значит, оно снизится и в тканях. При этом БФГ будет облегчать передачу кислорода от гемоглобина к тканям.
Некоторые вещества способны прочно связываться с гемоглобином или же вовсе менять его структуру. Одним из них является угарный газ, чье сродство к гемоглобину в 200 раз превышает таковое для кислорода. Отравления угарным газом часто происходят в помещениях с печным отоплением, при пожарах и авариях на производстве. Со временем кислород вытесняет угарный газ из гемоглобина, и в легких случаях пациенты помещаются под наблюдение и получают ингаляции с увлажненным кислородом. Необходимой мерой при тяжелых отравлениях угарным газом является переливание эритроцитарной массы.
К веществам, способным изменять структуру гемоглобина, относятся метгемоглобинобразователи — соединения, способные окислять двухвалентное железо в геме до трехвалентного. К ним относятся нитриты, нитраты, некоторые местные анестетики, аминофенолы, хлораты, примахин и некоторые сульфаниламиды. Состояние, характеризующееся появлением в крови окисленного гемоглобина, называют метгемоглобинемией. При высокой метгемоглобинемии капля крови, помещенная на фильтровальную бумагу, имеет характерный коричневый цвет, а при пропускании кислорода через пробирку с такой кровью ее цвет не меняется. Метгемоглобинемия выше 70 % от общего содержания гемоглобина часто приводит к гибели пациента еще до момента постановки диагноза.
Источники:
- Harrison’s hematology and oncology Longo, Dan L (Dan Louis), Third edition. New York : McGraw-Hill Education Medical, 2017.
- Наглядная физиология, С. Зильбернагль, А. Деспопулос, 2013.
- Ленинджер А. Основы биохимии: В 3-х т. Т. 1. /Д. Нельсон, М. Кокс ; Пер. с англ.-М.: БИНОМ: Лаборатория знаний, 2011.- 694 с.
Нашли опечатку? Выделите фрагмент и нажмите Ctrl+Enter.
Источник