Структура и свойства гемоглобина и миоглобина
Содержание статьи
МИОГЛОБИН
МИОГЛОБИН (myoglobinum; греч, mys, my [os] мышца + лат. globus шарик; Mgb; син.: миогемоглобин, мышечный гемоглобин) — сложный белок красного цвета, относящийся к хромопротеидам; содержится в красных мышцах животных различных классов и видов. Основной функцией М. является накопление кислорода, поступающего в процессе дыхания в мышцы из крови и отдача кислорода по мере необходимости в нем (см. Газообмен).
Факторами, определяющими различия в содержании М., являются видовая принадлежность животных (см. табл.), особенности условий их существования, образ жизни, тип мышцы, степень ее активности, а также трофические влияния нервной системы. Очень высокое содержание М. (до 16—40 г на 100 г высушенной ткани) характерно для водных животных. Значительные количества М. содержатся в красных мышцах нек-рых беспозвоночных (моллюски), в гладких мышцах мускульного желудка птиц, в гладкой мускулатуре бронхов, сосудов, стенок кишечника, а также в паренхиматозных органах, причем последние участвуют в процессах синтеза (печень), распада (селезенка) и выделения пигмента. Однако еще не доказано, что пигмент, выделенный из паренхиматозных органов, идентичен М.
Таблица. Содержание миоглобина в сердечных и скелетных мышцах человека и некоторых классов позвоночных животных (в г на 100 г высушенной ткани)
Класс животных; человек | Мышцы сердца | Скелетные мышцы |
Рыбы | 0,2—0,5 | 0, 1—0,4 |
Амфибии | 0, 1—0, 6 | 0 —0,1 |
Рептилии | 0, 8—2, 6 | 0,2—2,4 |
Птицы: | ||
домашние | 1, 0-2,0 | 0,9—1,8 |
дикие | 2,1—6, 5 | 0, 7—5, 3 |
Млекопитающие: | ||
домашние | 0, 9—4, 4 | 0,2-8, 5 |
дикие | 1,1-6,2 | 0,02—5,13 |
Человек | 1,5 | 1,4-3,9 |
В крови и моче М. в норме отсутствует. Поэтому его появление в этих биол, жидкостях является признаком заболеваний, сопряженных с деструкцией тканей (инфаркт миокарда, травматические повреждения мышц, генетически обусловленная прогрессивная мышечная дистрофия и т. д.). Определение уровня М. в плазме крови может служить критерием обширности деструкции, динамики процесса и эффективности лечебных мероприятий.
Мол. вес (масса) М. составляет в среднем 17 500, содержание железа 0,34%, Изоэлектрическая точка находится при pH 6,99. М. характеризуется большой растворимостью и высокой устойчивостью к действию щелочей.

Схематическое изображение молекулы миоглобина: цифрами отмечен каждый десятый аминокислотный остаток; светлые кружки — остатки пролина (Про), часто совпадающие с поворотом полипептидной цепи. В виде диска изображена группа гема; буквами С и N обозначены концевые участки цепи, содержащие соответственно свободные карбокси- и аминогруппы.
Изучена третичная структура М. и построена пространственная модель его молекулы (рис.). М. состоит из простого белка— глобина, представляющего собой одну полипептидную цепь, в состав к-рой входит 153 аминокислотных остатка, и простетической (небелковой) группы — гема (железопротопорфириновый комплекс, или протогем). Полипептидная цепь глобина М. на 77% представлена альфа-спиральными участками (всего 8 спиралей), к-рые перемежаются с участками неупорядоченной структуры, расположенными в области четырех изгибов полипептидной цепи. Все полярные группы, образованные лизином, аргинином, глутамином, аспарагином, гистидином, треонином, тирозином и триптофаном, находятся на поверхности и соединены с молекулами воды, а неполярные остатки сконцентрированы в центре. Поэтому молекула М. компактна и устойчива к изменениям pH и ионной силы р-ра.
По своим свойствам глобин М. близок к альбуминам. Гем М. расположен в углублении, находящемся на определенном изгибе полипептидной цепи вблизи поверхности молекулы. Это делает гем более доступным различным воздействиям. Одной координационной связью (пятой) гем соединен с гистидиновым остатком альфа-спирали, а шестая координационная связь железа гема может быть занята водой, кислородом, окисью углерода или другими соединениями. При взаимодействии с кислородом (оксигенации) в глобине молекулы М. не происходит структурных изменений. Пигмент мышц не способен соединяться с двуокисью углерода (CO2).
Доказано существование двух конформеров (А и В) нативного состояния М., к-рые отличаются по своим свойствам и структуре. Переход конформера А в конформер В протекает при повышении температуры от 20 до 40° и отражается на активности гема. Высокотемпературный конформер В является менее реакционноспособным.
М. человека и животных имеет неодинаковую форму кристаллов: тонкие, игловидные, собранные в пучки (человек), ромбические таблички (рогатый скот) и др. М. подобно гемоглобину (см.) образует различные производные, отличающиеся друг от друга по спектрам поглощения. При соединении с кислородом М. превращается в оксимиоглобин (MgbO2), в к-ром железо гема является двухвалентным. С помощью метода дифракции рентгеновских лучей показано, что оксигенация М. сопровождается пространственным смещением железа от середины порфиринового кольца по направлению к проксимальному гистидину (F-8) полипептидной цепи на 0,033 нм. В гемоглобине же оксигенация вызывает гораздо большее смещение атома железа (0,04— 0,05 нм) и изменения в четвертичной структуре всей молекулы пигмента крови.
Вдыхание животными воздуха, содержащего окись углерода в повышающихся концентрациях (0,01—0,2%), приводит не только к увеличению содержания карбоксигемоглобина в крови, но и к связыванию окиси углерода миоглобином с образованием карбоксимиоглобина (MgbCO), железо гема к-рого также находится в двухвалентном состоянии. Когда количество карбоксигемоглобина в крови достигает 60%, содержание карбоксимиоглобина в скелетных мышцах составляет 10—50%, а в мышце сердца — от 6 до 44%. MgbCO обнаружен и в мышцах людей, погибших от отравления: угарным газом. Воздействие на М. окислителей обусловливает возникновение метмиоглобина (метMgb). Превращение М. в метMgb при окислении происходит очень легко и при одинаковых условиях в 12—14 раз превышает скорость образовании метгемоглобина. В то же время в мышцах животных эта реакция после внутривенного вливания окисляющих ядов протекает более медленно, чем окисление гемоглобина.
Максимумы полос поглощения М. и его производных расположены при следующих значениях длин волн: для MgbO2 — 582, 542, 415 нм; для Mgb — 602, 560, 436 нм; для MgbCO — 585, 542, 423 нм; для метMgb — 630, 500, 409 нм. Известный другие производные пигмента: сульфмиоглобин, нитроксимиоглобин, цианмиоглобин и т. д. В реакциях сопряженного окисления М. переходит в зеленый пигмент — вердомиоглобин, а при более глубоком распаде — в желчные пигменты (см.).
Биосинтез М. происходит в мышцах со значительно меньшей скоростью по сравнению с гемоглобином. После введения животным меченого железа оно обнаруживается в гемоглобине уже спустя 6—8 дней, а в М. только через месяц. Продолжительность существования М. составляет 80 дней. В процессе эмбриогенеза М. раньше всего появляется в мышце сердца.
В мышцах позвоночных М. локализуется в саркоплазме на уровне диска А и связан электростатически с наружной мембраной митохондрий или саркоплазматического ретикулума (см. Мышечная ткань).
Создаваемый с помощью М. резерв кислорода в мышцах прежде всего определяется концентрацией в них пигмента и его способностью подвергаться оксигенации и деоксигенации. М. поддерживает постоянный уровень оксигенации во время мышечного сокращения, создавая определенный градиент напряжения кислорода между капиллярами и мышечными клетками, обеспечивая, т. о., возможность его утилизации при усилении окислительных процессов в работающих мышцах. Благодаря высокому сродству к кислороду полунасыщение им М. происходит менее чем за 0,1 сек. (при pO2 = 3,3 мм рт. ст.). При pO2, равном 5 мм рт. ст. (минимальное для деятельности цитохромоксидазы), диссоциирует до 40% оксимиоглобина, повышая, т. о., количество кислорода, растворенного в саркоплазме. М. участвует в молекулярном механизме его транспорта в клетку путем создания градиента и ускорения диффузии. Не исключена возможность непосредственного переноса кислорода М. при поступательном движении молекулы пигмента. Благодаря наличию пероксидазных свойств М. обладает ферментативной активностью. Доказана возможность включения М. в работу фосфорилирующей дыхательной цепи в качестве акцептора электронов от восстановленных коферментов пиридинового типа (НАД-H и в меньшей степени НАДФ-Н).
Существующие методы количественной оценки содержания М. включают электрофорез (см.) и ионообменную хроматографию (см.). Разделение М. и гемоглобина достигается благодаря их различной растворимости в 3 М фосфатном буфере при pH 6,6 (гемоглобин при этом значении pH нерастворим). Спектрофотометрическая регистрация количества М. основана на разнице в светопоглощении карбокси- и цианметсоединений М. и гемоглобина в видимой части спектра (см. Спектрофотометрия). Предложены также гистохимические методы определения М., основанные на бензидин-пероксидазной активности пигмента. Они позволяют с достаточной достоверностью выявлять М. в мышечной ткани и устанавливать закономерности в локализации и распределении М. в норме и их изменения при различных патол, состояниях.
Миоглобин в судебно-медицинском отношении
Миоглобин в судебно-медицинском отношении имеет значение в случаях прижизненного поражения скелетных мышц, сопровождающегося высвобождением М. При этом М. накапливается в плазме крови. При концентрации, превышающей 30 мг%, развивается миоглобинурия (см.), к-рая наряду с шоковым и коматозным состоянием, интоксикацией и пр. служит патогенетическим фактором развития миоглобинурийного нефроза (см. Нефротический синдром). Повреждение мышц, сочетающееся с обширными гематомами или внутрисосудистым гемолизом, кроме того, ведет к возникновению гемоглобинемии (см.), приводящей в последующем к миоглобинурии и миоглобинурийному нефрозу. Т. о., миоглобинемия и обусловленные ею процессы служат экспертным критерием поражения скелетной мускулатуры и доказательством его прижизненности.
Миоглобинемия может иметь место при механической травме (одномоментная травма обширных мышечных массивов, так наз. краш-синдром, или синдром раздавливания), отравлении миолитическими ядами, в т. ч. пищевыми (токсический миозит), при нарушении артериального или венозного кровообращения в конечностях (тромбоз или тромбоэмболия крупных артерий, тромбоз вен, синдром жгута, состояние после реплантации конечностей), при длительном пребывании человека в одной и той же позе (позиционное сдавление), при ожогах, обморожениях, при судорожных состояниях (эпилепсия, столбняк и т. д.).
Наличие и содержание М. в крови и моче устанавливают спектрофотометрическим способом или при помощи электрофореза. Для дифференциации пигментов в моче применяют пробу с сульфатом аммония, при к-рой в осадке мочи, содержащем миоглобиновые шлаки, на фильтре наблюдается характерное окрашивание .
Библиография: Атанасов Б. П. Модели конформеров нативного состояния миоглобина, Молек. биол., т. 4, в. 3, с. 348, 1970, библиогр. ; Верболович П. А. и Верболович В. П. Миоглобин и использование кислорода в животном организме, в кн.: Полярографическое определение кислорода в биол, объектах, под ред. В.А. Березовского, с. 123, Киев, 1974, библиогр.; Свадковский Б. С. Острый пигментный нефроз и его судебно-медицинская оценка, М., 1974, библиогр.; Троицкая О. В. Миоглобин, его химическое строение и функции в организме, Вопр, мед. хим., т. 17, в. 5, с. 451, 1971, библиогр.; Murray J. D. On the role of myoglobin in muscle respiration, J. theor. Biol., v. 47, p. 115, 1974, bibliogr.; Wittenberg J. B. Myoglobin — facilitated oxygen diffusion, role of myoglobin in oxygen entry into muscle, Physiol. Rev., v. 50, p. 559, 1970, bibliogr.
П. А. Верболович, В. П. Верболович; М. В. Кисин (суд.-мед.).
Источник
Структура и функции сложных белков. Строение и функции миоглобина и гемоглобина.
Тема: «СТРОЕНИЕ И ФУНКЦИИ СЛОЖНЫХ БЕЛКОВ. СТРУКТУРА И ФУНКЦИИ МИОГЛОБИНА И ГЕМОГЛОБИНА»
1. Определение понятия и основные принципы классификации сложных белков. Примеры.
2. Гликопротеины: особенности строения, примеры, функции. Иммуноглобулины: особенности строения, классы иммуноглобулинов, их роль в организме.
3. Фосфопротеины: представители, связывание простетической группы с апопротеином, роль в организме.
4. Металлопротеины: представители, характер простетической группы, ее связывание с апопротеином, роль металлопротеинов в организме.
5. Строение и функции хромопротеинов (на примере миоглобина и гемоглобина). Аллостерические свойства гемоглобина. Кооперативный эффект. Роль СО2, водородных ионов и 2,3-дифосфоглицерата в регуляции сродства гемоглобина к кислороду.
6. Производные гемоглобина (дезоксигемоглобин, оксигемоглобин, карбгемоглобин, карбоксигемоглобин, метгемоглобин, цианметгемоглобин), их характеристика. Причины и последствия накопления метгемоглобина в крови.
7. Молекулярные формы гемоглобина. Фетальный гемоглобин, особенности структуры, свойства, биологическая роль. Серповидно-клеточный гемоглобин (HbS), особенности структуры, свойства, проявления гемоглобиноза S.
| Раздел 3.1 | Понятие о простых и сложных белках. Классификация сложных белков. | |||||||||||||||||||||
3.1.1. Запомните, что белки можно разделить на простые и сложные. Простые белки — белки, которые состоят только из аминокислотных остатков. Сложные белки (холопротеины) — белки, которые состоят из аминокислотных остатков и компонентов небелковой природы. Белковая часть сложного белка получила название апопротеин, небелковая часть сложного белка — простетическая группа. 3.1.2. Сложные белки классифицируют в зависимости от химического строения их простетических групп. Выучите названия классов сложных белков, их простетических групп и примеры представителей различных классов сложных белков (таблица 3.1) .
В этом разделе мы более подробно коснёмся строения и свойств хромопротеинов и нуклеопротеинов. Характеристика белков — представителей других классов будет рассмотрена позднее. | ||||||||||||||||||||||
| Раздел 3.2 | Особенности строения и биологическая роль липопротеинов, нуклеопротеинов, фосфопротеинов, металлопротеинов. |
3.2.1. Липопротеины – сложные белки, содержащие в качестве простетической группы липиды и их производные. Представителями служат хиломикроны и другие фракции липопротеинов крови. Липопротеины являются транспортными формами липидов в крови. Более подробно этот класс сложных белков рассматривается в разделе «Обмен липидов». 3.2.2. Нуклеопротеины – сложные белки, содержащие в качестве простетической группы нуклеиновые кислоты. Нуклеопротеинами являются вирусы, рибосомы, хроматин клеточного ядра. Существуют две разновидности нуклеопротеинов: 1) рибонуклеопротеины, в состав которых входит рибонуклеиновая кислота (РНК); 2) дезоксирибонуклеопротеины, в состав которых входит дезоксирибонуклеиновая кислота (ДНК). Строение и свойства нуклеиновых кислот будут рассматриваться позднее. Особенность белковой части нуклеопротеинов заключается в том, что в её состав входит много положительно заряженных аминокислотных остатков. Так, в состав дезоксирибонуклеопротеинов входят белки гистоны, богатые лизином и аргинином. Между белковыми и небелковыми компонентами нуклеопротеинов образуются ионные связи (так как нуклеиновые кислоты заряжены отрицательно). 3.2.3. Фосфопротеины содержат в своём составе остатки фосфорной кислоты, соединённые с остатками гидроксиаминокислот (серин, треонин) при помощи сложноэфирных связей. К этой группе сложных белков относятся казеиноген молока, яичные белки овальбумин и вителлин. Многие внутриклеточные белки являются фосфопротеинами. Присоединение фосфатной группы к белку часто вызывает изменение его функции. Фосфорилирование и обратный процесс – дефосфорилирование – распространенный механизм регуляции биологической активности белков. Например, фосфорилирование гистонов снижает их способность связываться с ДНК и участвовать в регуляции матричных синтезов с участием ДНК. 3.2.4. Металлопротеины. Если в белке содержатся ионы одного или нескольких металлов, то такие белки называются металлопротеинами. Ионы металлов соединены координационными связями с функциональными группами белка и участвуют в поддержании его пространственной структуры. Металлопротеины часто являются ферментами. Трансферрин – водорастворимый железопротеин, содержащийся в сыворотке крови в составе β-глобулинов. Молекула трансферрина содержит 2 иона Fe3+; этот белок служит переносчиком железа в организме. Ферритин – внутриклеточный глобулярный белок, содержится главным образом в селезенке, печени, костном мозге, выполняя роль депо железа в организме. Благодаря ферритину цитозольные запасы железа поддерживаются в растворимой и нетоксичной форме. Гемосидерин, в отличие от ферритина и трансферрина, является водонерастворимым железосодержащим белковым комплексом. Он содержится главным образом в клетках печени и селезенки, накапливается при избытке железа в организме, например, при частых переливаниях крови. Церулоплазмин – белок α2-глобулиновой фракции сыворотки крови, его молекула содержит 6 – 8 ионов меди. Обладает каталитической активностью, катализирует реакцию окисления Fe2+ в Fe3+. Это делает возможным связывание железа с трансферрином и его последующий транспорт в крови. | |
| Раздел 3.3 | Строение и функции гликопротеинов. Иммуноглобулины. |
3.3.1. Гликопротеины – содержат в качестве простетической группы углеводы и их производные. Они присоединяются либо N-гликозидной связью к амидогруппе остатка аспарагина, либо О-гликозидной связью к гидроксигруппе остатка серина или треонина. Углеводная часть имеет нерегулярное строение. Гликопротеины выполняют в организме следующие функции: структурную (коллаген, эластин), защитную (антитела, интерфероны), рецепторную, гормональную (гормоны гипофиза), ферментативную, транспортную. 3.3.2. Иммуноглобулины (антитела) — группа белков, вырабатываемых организмом в ответ на попадание в организм чужеродных структур (антигенов). Они синтезируются В-лимфоцитами или плазматическими клетками. В организме может вырабатываться порядка 107 разновидностей иммуноглобулинов, каждый из которых может распознавать определённый антиген. Все иммуноглобулины подразделяют на пять классов: IgA, IgG, IgM, IgD, IgE. Основную структурную единицу иммуноглобулинов, или мономер, образуют четыре полипептидные цепи, соединённые между собой дисульфидными связями, из них: а) две идентичные тяжёлые цепи (молекулярная масса 53000 — 75000 Да), обозначаемые буквами Н; б) две идентичные лёгкие цепи (молекулярная масса около 23000 Да), обозначаемые буквами L. Иммуноглобулины G, D и Е по своей структуре, как правило, являются мономерами, молекулы IgM построены из пяти мономеров, IgA могут быть как мономерами, так и состоять из двух и более структурных единиц. Белковые цепи, входящие в состав иммуноглобулинов, можно условно разделить на специфические домены, или области, имеющие определённые структурные и функциональные особенности. N-концевые участки как L-, так и Н-цепей называются вариабельной областью (V), так как их структура характеризуется существенными различиями у разных классов антител. Внутри вариабельного домена имеются 3 гипервариабельных участка, отличающихся наибольшим разнообразием аминокислотной последовательности. Именно вариабельная область антител ответственна за связывание антигенов по принципу комплементарности; первичная структура белковых цепей в этой области определяет специфичность антител. | |
С-концевые домены Н- и L-цепей обладают относительно постоянной первичной структурой в пределах каждого класса антител и называются константной областью (С). Константная область определяет свойства различных классов иммуноглобулинов, их распределение в организме, может принимать участие в запуске механизмов, вызывающих уничтожение антигенов.
3.3.3. Функции иммуноглобулинов. IgG составляют около 75% общего количества иммуноглобулинов плазмы крови. IgG эффективно связывают и инактивируют чужеродные молекулы и клетки, попавшие в организм, а также облегчают их дальнейшее уничтожение, способны преодолевать плацентарный барьер, что обеспечивает иммунитет новорождённых в течение первых недель жизни.
IgA содержатся главным образом в секретах слизистых оболочек дыхательных и выделительных путей, желудочно-кишечного тракта, т.е. обеспечивают защиту поверхностей, сообщающихся с внешней средой.
IgM синтезируются на ранних стадиях иммунного ответа, вступают в реакцию агглютинации с антигенами, активируют систему комплемента.
IgD связаны с мембраной лимфоцитов, функционируют в качестве рецепторов для антигенов.
IgE участвуют в развитии аллергических реакций, в защите от паразитарных инвазий.
| Раздел 3.4 | Миоглобин и гемоглобин: сходство и различия структуры и функции. |
| 3.4.1. Среди хромопротеинов различают гемопротеины (содержат в качестве простетической группы порфириновые производные) и флавопротеины (содержат производные рибофлавина — витамина B2). Хромопротеины участвуют в осуществлении многих жизненно важных функций, таких как тканевое дыхание, перенос кислорода, окислительно-восстановительные реакции, светоощущение, фотосинтез в растительных клетках и другие процессы. 3.4.2. К гемопротеинам относятся: гемоглобин, миоглобин, цитохромы, пероксидаза, каталаза. Эти белки содержат в качестве простетической группой гем.
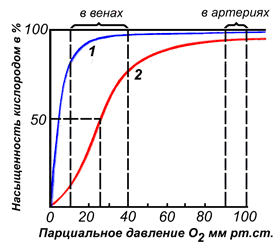
По своему химическому строению гем представляет собой протопорфирин IX, связанный с двухвалентным железом. Протопорфирин IX — органическое соединение, относящееся к классу порфиринов. Протопорфирин IX содержит четыре замещённых пиррольных кольца, соединённых метиновыми мостиками =СН—. Заместителями в пиррольных кольцах являются: четыре метильные группы СН3—, две винильные группы СН2=СН— и два остатка пропионовой кислоты — СН2—СН2—СООН. Гем соединяется с белковой частью следующим образом. Неполярные группы . протопорфирина IX взаимодействуют с гидрофобными участками аминокислот при помощи гидрофобных связей. Кроме того, имеется координационная связь между атомом железа и имидазольным радикалом гистидина в белковой цепи. Ещё одна координационная связь атома железа может использоваться для связывания кислорода и других лигандов. Присутствие в биологическом материале гемсодержащих белков обнаруживается при помощи бензидиновой пробы (при добавлении бензидина и пероксида водорода исследуемый раствор окрашивается в сине-зелёный цвет). 3.4.3. Сравните структуру и функцию миоглобина и гемоглобина, запомните характерные особенности каждого из этих белков. Миоглобин — хромопротеин, присутствующий в мышечной ткани и обладающий большим сродством к кислороду. Молекулярная масса этого белка около 16000 Да, Молекула миоглобина имеет третичную структуру и представляет собой одну полипептидную цепь, соединённую с гемом. Миоглобин не обладает аллостерическими свойствами (см. 2.4.), кривая насыщения его кислородом имеет вид гиперболы (рисунок 4). Функция миоглобина заключается в создании в мышцах кислородного резерва, который расходуется по мере необходимости, восполняя временную нехватку кислорода. Гемоглобин (Hb) — хромопротеин, присутствующий в эритроцитах и участвующий в транспорте кислорода к тканям. Гемоглобин взрослых людей называется гемоглобином А (Hb A). Молекулярная масса его составляет около 65000 Да. Молекула Hb А имеет четвертичную структуру и включает четыре субъединицы — полипептидные цепи (обозначаемые α1, α2, β1 и β2, каждая из которых связана с гемом. Запомните, что гемоглобин относится к аллостерическим белкам, его молекулы могут обратимо переходить из одной конформации в другую. При этом изменяется сродство белка к лигандам. Конформация, обладающая наименьшим сродством к лиганду, называется напряжённой, или Т-конформацией. Конформация, обладающая наибольшим сродством к лиганду, называется релаксированной, или R-конформацией. R- и Т-конформации молекулы гемоглобина находятся в состоянии динамического равновесия: Различные факторы среды могут сдвигать это равновесие в ту или иную сторону. Аллостерическими регуляторами, влияющими на сродство Hb к O2, являются: 1) кислород; 2) концентрация Н+ (рН среды); 3) углекислота (СO2) ; 4) 2,3-дифосфоглицерат (ДФГ) . Присоединение молекулы кислорода к одной из субъединиц гемоглобина способствует переходу напряжённой конформации в релаксированную и повышает сродство к кислороду других субъединиц той же молекулы гемоглобина. Это явление получило название кооперативного эффекта. Сложный характер связывания гемоглобина с кислородом отражает кривая насыщения гемоглобина O2, имеющая S-образную форму (рисунок 3.1). Повышение содержания СO2, Н+, ДФГ на фоне низкого парциального давления O2 в тканях способствует взаимодействию этих факторов с гемоглобином и переходу R-конформации в Т-конформацию. Это приводит к смещению равновесия в уравнении (1) вправо. Выделившийся O2 поступает в ткани.
Рисунок 3.1. Кривые насыщения миоглобина (1) и гемоглобина (2) кислородом. | |
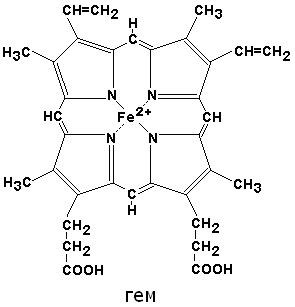
| Раздел 3.5 | Производные гемоглобина. Молекулярные формы гемоглобина: отличия HbF и HbS от HbA. |
3.5.1. Следует различать производные гемоглобина и его молекулярные формы. К производным гемоглобина относятся продукты взаимодействия гемоглобина (дезоксигемоглобина) с различными лигандами. Это взаимодействие, как правило, носит обратимый характер. Производные гемоглобина, представляющие наибольший интерес для медицины, перечислены в таблице 3.2. Запомните, что метгемоглобин (MetHb), в отличие от гемоглобина, теряет способность связывать и транспортировать кислород. Следовательно, попадание в организм больших количеств веществ, вызывающих образование метгемоглобина (нитриты, нитраты, анилин, нитробензол, некоторые лекарства), может привести к гипоксии (кислородному голоданию) тканей и смерти. В то же время метгемоглобин может легко связывать ионы CN—, нейтрализуя их токсическое действие. В результате образуется цианметгемоглобин. 3.5.2. Молекулярные формы гемоглобина отличаются друг от друга строением полипептидных цепей. Примером такой разновидности гемоглобина, существующей в физиологических условиях, является фетальный гемоглобин (HbF), присутствующий в крови в эмбриональной стадии развития человека. В отличие от HbA, его молекула содержит 2 α- и 2 γ-цепи (то есть β-цепи заменены на γ-цепи). Такой гемоглобин обладает более высоким сродством к кислороду. Именно это позволяет эмбриону получать кислород из крови матери через плаценту. Вскоре после рождения HbF в крови ребёнка замещается на HbA. В качестве примера аномального или патологического гемоглобина можно привести уже упоминавшийся (см. 2.4.) гемоглобин S, обнаруженный у больных серповидно-клеточной анемией. Как вам уже известно, он отличается от гемоглобина А заменой в β-цепях глутамата на валин. Эта аминокислотная замена вызывает снижение растворимости HbS в воде и уменьшение его сродства к O2. | |
| Критерий | Дезоксигемоглобин | Оксигемоглобин | Карбгемоглобин | Карбоксигемоглобин | Метгемоглобин | Цианметгемоглобин |
|---|---|---|---|---|---|---|
| Лиганд | — | O2 | СО2 | СО | ОН— | CN— |
| Валентность железа | II | II | II | II | III | III |
| Место присоединения лиганда | — | Fe2+ | NH2- группы глобина | Fe2+ | Fe3+ | Fe3+ |
| Механизм образования | — | Hb + O2 « HbO2 | Hb-NH2 + CO2 « Hb-NH-COOH | HbО2 + CO « HbCO + O2 | Hb + O2 + OН— « MetHb(OH) + O2— | MetHb(OH) + CN— « MetHb(CN) + OH— |
| Сродство к O2 | низкое | высокое | низкое | очень низкое | отсутствует | отсутствует |
| Присутствие в крови в норме | присутствует | присутствует | присутствует | отсутствует | отсутствует | отсутствует |
| Примеры | Обучающие задачи и эталоны их решения |
3.5.1. Задачи.1. В гидролизате сложного белка обнаружена фосфорная кислота. Можно ли с достаточной уверенностью сказать, к какому классу относится изучаемый белок? 2. Сравните растворимость простетической группы гемоглобина в воде и органических растворителях. 3. У альпиниста через 2 дня после подъёма на высоту 4000 м над уровнем моря содержание 2,3-дифосфоглицерата в эритроцитах составляет 6,5 ммоль/л (в норме — 4,5 ммоль/л) . Как это отразится на сродстве гемоглобина к кислороду? Какое значение для организма имеет такое изменение? 3.5.2. Эталоны решения.1. Как видно из таблицы 3.1, фосфорная кислота является простетической группой белков класса фосфопротеинов. Известно также, что фосфат входит в состав нуклеиновых кислот, которые могут выступать в качестве простетической группы нуклеопротеинов. Поэтому фосфорная кислота может быть обнаружена в гидролизате как фосфопротеина, так и нуклеопротеина. Для уточнения класса гидролизованного белка нужно провести с гидролизатом качественные реакции на азотистые основания и пентозы (см. 3.1.). 2. Растворимость вещества в воде или органических растворителях зависит от того, какие функциональные группы преобладают в молекуле. Простетическая группа гемоглобина — гем — содержит только две полярные группировки (две карбоксильные группы), остальная часть молекулы неполярна (равномерное распределение электронной плотности). Поэтому гем хорошо растворяется в органических растворителях и не растворяется в воде (см. 3.2.). 3. 2, 3-Дифосфоглицерат (ДФГ) является аллостерическим регулятором, понижающим сродство гемоглобина к кислороду. Поэтому при увеличении содержания ДФГ в эритроцитах равновесие в уравнении диссоциации HbO2 смещается вправо. Увеличение отдачи кислорода оксигемоглобином имеет компенсаторное значение для организма, так как восполняет недостаточное поступление кислорода из атмосферы (см. 3.2.). | |
Источник