Транспорт холестерина из печени
Содержание статьи
ОБМЕН, РОЛЬ И ТРАНСПОРТ ХОЛЕСТЕРИНА — Студопедия
В сутки в организме синтезируется около 1 г холестерина (рис.10). Основное место синтеза – печень (до 80%), меньше синтезируется в кишечнике, коже и других тканях. С пищей поступает около 0,4 г холестерина, его источником является только пища животного происхождения. Холестерин необходим для построения всех мембран, в печени из него синтезируются желчные кислоты, в эндокринных железах – стероидные гормоны, в коже – витамин Д.
Рис.10 Холестерин
Сложный путь синтеза холестерина можно поделить на 3 этапа (рис.11). Первый этап заканчивается образованием мевалоновой кислоты. Источником для синтеза холестерина служит ацетил-КоА. Сначала из 3 молекул ацетил-КоА образуется ГМГ-КоА – общий предшественник в синтезе холестерина и кетоновых тел (однако реакции синтеза кетоновых тел происходят в митохондриях печени, а реакции синтеза холестерина – в цитозоле клеток). Затем ГМГ-КоА под действием ГМГ-КоА-редуктазы восстанавливается до мевалоновой кислоты с использованием 2 молекул НАДФН. Эта реакция является регуляторной в синтезе холестерина. Синтез холестерина тормозит сам холестерин, желчные кислоты и гормон голода глюкагон. Усиливается синтез холестерина при стрессе катехоламинами.
На втором этапе синтеза из 6 молекул мевалоновой кислоты образуется углеводород сквален, имеющий линейную структуру и состоящий из 30 атомов углерода.
На третьем этапе синтеза происходит циклизация углеводородной цепи и отщепление 3 атомов углерода, поэтому холестерин содержит 27 углеродных атомов. Холестерин является гидрофобной молекулой, поэтому транспортируется кровью только в составе разных липопротеинов.
Рис. 11 Синтез холестерина
Липопротеины – липид-белковые комплексы, предназначенные для транспорта нерастворимых в водных средах липидов по крови (рис.12). Снаружи липопротеины (ЛП) имеют гидрофильную оболочку, которая состоит из молекул белков и гидрофильных групп фосфолипидов. Внутри ЛП находятся гидрофобные части фосфолипидов, нерастворимые молекулы холестерина, его эфиров, молекулы жиров. ЛП делятся (по плотности и подвижности в электрическом поле) на 4 класса. Плотность ЛП определяется соотношением белков и липидов. Чем больше белка, тем больше плотность и тем меньше размер.
Рис.12. Строение липопротеидов
· 1 класс – хиломикроны (ХМ). Содержат 2% белка и 98% липидов, среди липидов преобладают экзогенные жиры, переносят экзогенные жиры от кишечника к органам и тканям, синтезируются в кишечнике, в крови присутствуют непостоянно – только после переваривания и всасывания жирной пищи.
· 2 класс – ЛП очень низкой плотности (ЛПОНП) или пре-b-ЛП. Белка в них 10%, липидов – 90%, среди липидов преобладают эндогенные жиры, транспортируют эндогенные жиры из печени в жировую ткань. Основное место синтеза – печень, небольшой вклад вносит тонкий кишечник.
· 3 класс – ЛП низкой плотности (ЛПНП) или b-ЛП. Белка в них 22% , липидов – 78%, среди липидов преобладает холестерин. Нагружают клетки холестерином, поэтому их называют атерогенными, т.е. способствующими развитию атеросклероза (АС). Образуются непосредственно в плазме крови из ЛПОНП под действием фермента ЛП-липазы.
· 4 класс ЛП высокой плотности (ЛПВП) или a-ЛП. Белка и липидов содержат по 50%, среди липидов преобладают фосфолипиды и холестерин. Разгружают клетки от избытка холестерина, поэтому являются антиатерогенными, т.е. препятствующими развитию АС. Основное место их синтеза – печень, небольшой вклад вносит тонкий кишечник.
Транспорт холестерина липопротеинами.
Печень является основнымместом синтеза холестерина. Холестерин, синтезированный в печени, упаковывается в ЛПОНП и в их составе секретируется в кровь. В крови на них действует ЛП-липаза, под влиянием которой ЛПОНП переходят в ЛПНП. Таким образом, ЛПНП становятся основной транспортной формой холестерина, в которой он доставляется к тканям. ЛПНП могут попадать в клетки двумя путями: рецепторным и нерецепторным захватом. Большинство клеток на своей поверхности имеют рецепторы к ЛПНП. Образовавшийся комплекс рецептор-ЛПНП эндоцитозом попадает внутрь клетки, где распадается на рецептор и ЛПНП. Из ЛПНП при участии лизосомальных ферментов освобождается холестерин. Этот холестерин используется для обновления мембран, тормозит синтез холестерина данной клеткой, а также, если количество холестерина, поступающего в клетку, превышает ее потребность, то подавляется и синтез рецепторов к ЛПНП.
Это уменьшает поток холестерина из крови в клетки, таким образом, клетки, для которых характерен рецепторный захват ЛПНП, имеют механизм, который ограждает их от избытка холестерина. Для гладкомышечных клеток сосудов и макрофагов характерен нерецепторный захват ЛПНП из крови. В эти клетки ЛПНП, а значит, и холестерин попадают диффузно, то есть, чем их больше в крови, тем больше их попадает в эти клетки. Эти разновидности клеток не имеют механизма, который ограждал бы их от избытка холестерина. В «обратном транспорте холестерина» от клеток участвуют ЛПВП. Они забирают избыток холестерина из клеток и возвращают его обратно в печень. Холестерин выводится с калом в виде желчных кислот, часть холестерина в составе желчи попадает в кишечник и также выводится с калом.
Источник
Холестерол используется как переносчик полиненасыщенных жирных кислот
Транспорт холестерола и его эфиров осуществляется липопротеинами низкой и высокой плотности.
Липопротеины высокой плотности
Общая характеристика
- образуются в печени de novo, в плазме крови при распаде хиломикронов, некоторое количество в стенке кишечника,
- в составе частицы примерно половину занимают белки, еще четверть фосфолипиды, остальное холестерин и ТАГ (50% белка, 25% ФЛ, 13% эфиров ХС и 5% свободного ХС, 7% ТАГ),
- структурным апобелком является апо А1, также содержат апоЕ и апоСII.
Функция
- Транспорт свободного ХС от тканей к печени.
- Фосфолипиды ЛПВП являются источником полиеновых кислот для синтеза клеточных фосфолипидов и эйкозаноидов.
Метаболизм
1. Синтезированные в печени частицы (насцентные или первичные ЛПВП) содержат в основном фосфолипиды и апобелки. Остальные липидные компоненты накапливаются в ЛПВП по мере метаболизма в плазме крови.
Новосинтезированные ЛПВП выглядят на электронных микрофотографиях как двухслойные диски из двух монослоев фосфолипидов (таблеткообразная форма). Синтез апоЕ и апоС происходит главным образом в печени, в отличие от апо А-I, образующегося как в печени, так и в тонком кишечнике.
2-3. В плазме крови насцентный ЛПВП сначала превращается в ЛПВП3 (условно его можно назвать «зрелый»). В этом превращении главным является то, что ЛПВП
- забирает от клеточных мембран свободный холестерин при непосредственном контакте или при участии специфических транспортных белков,
- взаимодействуя с мембранами клеток, отдает им часть фосфолипидов из своей оболочки, доставляя таким образом полиеновые жирные кислоты в клетки,
- тесно взаимодействует с ЛПНП и ЛПОНП, получая от них свободный холестерин. В обмен ЛПВП3 отдают эфиры ХС, образованные благодаря переносу жирной кислоты от фосфатидилхолина (ФХ) на холестерин (ЛХАТ-реакция, см п.4).
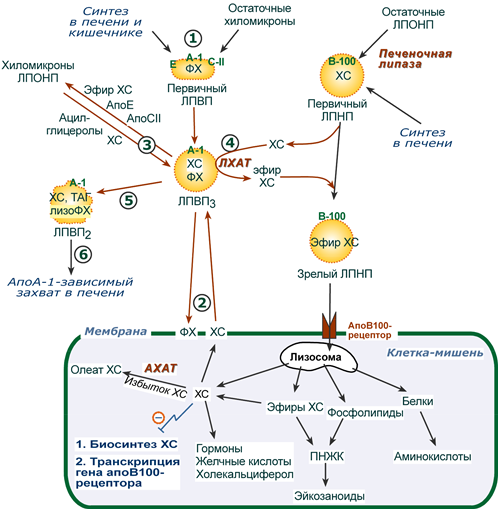
Транспорт холестерола и его эфиров в организме
(цифры соответствуют пунктам метаболизма ЛПВП по тексту)
4. Внутри ЛПВП активно протекает реакция при участии лецитин:холестерол-ацилтрансферазы (ЛХАТ-реакция). В этой реакции остаток полиненасыщенной жирной кислоты переносится от 2-го положения фосфатидилхолина (из оболочки самого ЛПВП) на получаемый свободный холестерин с образованием лизофосфатидилхолина (лизоФХ) и эфиров ХС. ЛизоФХ остается внутри ЛПВП, эфир холестерина отправляется в ЛПНП.
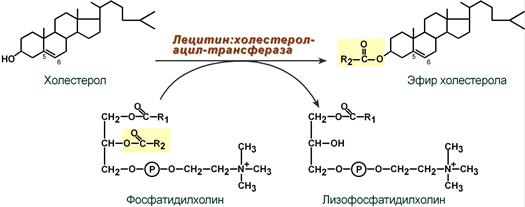
Реакция этерификации холестерола
при участии лецитин:холестерол-ацилтрансферазы
5. В результате первичный ЛПВП постепенно, через зрелую форму ЛПВП3, преобразуется в ЛПВП2 (остаточный, ремнантный). При этом происходят и дополнительные события:
- взаимодействуя с разными формами ЛПОНП и ХМ, ЛПВП получают ацил-глицеролы (МАГ, ДАГ, ТАГ), и обмениваются холестерином и его эфирами,
- ЛПВП отдают апоЕ- и апоСII-белки на первичные формы ЛПОНП и ХМ, и потом забирают обратно апоСII-белки от остаточных форм.
Таким образом, при метаболизме ЛПВП в нем происходит накопление свободного ХС, МАГ, ДАГ, ТАГ, лизоФХ и утрата фосфолипидной оболочки. Функциональные способности ЛПВП снижаются.
6. Далее ЛПВП2 захватывается гепатоцитами при помощи апоА-1-рецептора, происходит эндоцитоз и частица разрушается.
Липопротеины низкой плотности
Общая характеристика
- образуются в гепатоцитах de novo и в сосудистой системе печени под воздействием печеночной ТАГ-липазы из ЛПОНП,
- в составе преобладают холестерол и его эфиры, другую половину массы делят белки и фосфолипиды (38% эфиров ХС, 8% свободного ХС, 25% белки, 22% фосфолипидов, 7% триацилглицеролов),
- основным апобелком является апоВ-100,
- нормальное содержание в крови 3,2-4,5 г/л,
- самые атерогенные.
Функция
1. Транспорт холестерола в клетки, использующих его
- для реакций синтеза половых гормонов (половые железы), глюкокортикоидов и минералокортикоидов (кора надпочечников),
- для превращения в холекальциферол (кожа),
- для образования желчных кислот (печень),
- для выведения в составе желчи (печень).
2. Транспорт полиеновых жирных кислот в виде эфиров ХС в некоторые клетки рыхлой соединительной ткани (фибробласты, тромбоциты, эндотелий, гладкомышечные клетки), в эпителий гломерулярной мембраны почек, в клетки костного мозга, в клетки роговицы глаз, в нейроциты, в базофилы аденогипофиза.
Клетки рыхлой соединительной ткани активно синтезируют эйкозаноиды. Поэтому им необходим постоянный приток полиненасыщенных жирных кислот (ПНЖК), что осуществляется через апо-В-100-рецептор, т.е. регулируемым поглощением ЛПНП, которые несут ПНЖК в составе эфиров холестерола.
Особенностью клеток, поглощающих ЛПНП, является наличие лизосомальных кислых гидролаз, расщепляющих эфиры ХС. У других клеток таких ферментов нет.
Иллюстрацией значимости транспорта ПНЖК в указанные клетки служит ингибирование салицилатами фермента циклооксигеназы, образующей эйкозаноиды из ПНЖК. Салицилаты успешно применяются в кардиологии для подавления синтеза тромбоксанов и снижения тромбообразования, при лихорадке, как жаропонижающее средство за счет расслабления гладких мышц сосудов кожи и повышения теплоотдачи. Однако одним из побочных эффектов тех же салицилатов является подавление синтеза простагландинов в почках и снижение почечного кровобращения.
Также в мембраны всех клеток, как сказано выше (см «Метаболизм ЛПВП»), ПНЖК могут переходить в составе фосфолипидов от оболочки ЛПВП.
Метаболизм
1. В крови первичные ЛПНП взаимодействуют с ЛПВП, отдавая свободный ХС и получая этерифицированный. В результате в них происходит накопление эфиров ХС, увеличение гидрофобного ядра и «выталкивание» белка апоВ-100 на поверхность частицы. Таким образом, первичный ЛПНП переходит в зрелый.
2. На всех клетках, использующих ЛПНП, имеется высокоафинный рецептор, специфичный к ЛПНП – апоВ-100-рецептор. Около 50% ЛПНП взаимодействует с апоВ-100-рецепторами разных тканей и примерно столько же поглощается гепатоцитами.
3. При взаимодействии ЛПНП с рецептором происходит эндоцитоз липопротеина и его лизосомальный распад на составные части – фосфолипиды, белки (и далее до аминокислот), глицерол, жирные кислоты, холестерол и его эфиры.
- ХС превращается в гормоны или включается в состав мембран,
- излишки мембранного ХС удаляются с помощью ЛПВП,
- принесенные с эфирами ХС ПНЖК используются для синтеза эйкозаноидов или фосфолипидов.
- при невозможности удалить ХС часть его этерифицируется с олеиновой или линолевой кислотами ферментом ацил-SКоА:холестерол-ацилтрансферазой (АХАТ-реакция),
Синтез олеата холестерола при участии
ацил-SKoA-холестерол-ацилтрансферазы
На количество апоВ-100-рецепторов влияют гормоны:
- инсулин, тиреоидные и половые гормоны стимулируют синтез этих рецепторов,
- глюкокортикоиды уменьшают их количество.
Источник
ути транспорта холестерина
купить канцтовары в интернет магазине | В кровотоке переносчиками липидов являются липопротеины. Они состоят из липидного ядра, окруженного растворимыми фосфолипидами и свободным холестерином, а также апопротеинами, которые отвечают за направление липопротеинов к специфическим органам и тканевым рецепторам. Известно пять основных классов липопротеинов, различающихся по плотности, липидному составу и аполипопротеинам (табл. 5.1). Рис. 5.7 дает характеристику основных путей метаболизма циркулирующих липопротеинов. Жиры, поступившие с пищей, включаются в цикл, известный как экзогенный путь. Пищевые холестерин и триглицериды всасываются в кишечнике, включаются в хиломикроны клетками кишечного эпителия и транспортируются через лимфатические протоки в венозную систему. Эти большие, богатые триглицеридами частицы гидроли-зируются ферментом липопротеинлипазой, которая высвобождает жирные кислоты, захватываемые периферическими тканями, такими как жировая и мышечная. Образующиеся остатки хиломикронов состоят преимущественно из холестерина. Эти остатки поглощаются печенью, которая затем выделяет липиды в виде свободного холестерина либо желчных кислот обратно в кишечник. Эндогенный путь начинается с того, что липопротеины очень низкой плотности (ЛОНП) высвобождаются из печени в кровоток. Хотя основным липидным компонентом ЛОНП являются триглицериды, содержащие мало холестерина, основная часть холестерина поступает из печени в кровь именно в составе ЛОНП. Рис. 5.7. Обзор системы транспорта липопротеинов. Экзогенный путь: в желудочно-кишечном тракте пищевые жиры включаются в хиломикроны и через лимфатическую систему попадают в циркулирующую кровь. Свободные жирные кислоты (СЖК) поглощаются периферическими клетками (например, жировой и мышечной тканью); остатки (ремнанты) липопротеинов возвращаются в печень, где их холестериновая составляющая может транспортироваться обратно в ЖК тракт или использоваться в других метаболических процессах. Эндогенный путь: в печени синтезируются и поступают в кровь богатые триглицеридами липопротеины очень низкой плотности (ЛОНП), и их СЖК поглощаются и накапливаются в периферических жировых клетках и мышцах. Образующиеся в результате липопротеины промежуточной плотности (ЛПП) превращаются в липопротеины низкой плотности, основной циркулирующий липопротеин, осуществляющий транспорт холестерина. Большая часть ЛНП захватывается печенью и другими периферическими клетками путем рецептор-опосредованного эндоцитоза. Обратный транспорт холестерина, освобождаемого периферическими клетками, осуществляется липопротеинами высокой плотности (ЛВП), которые превращаются в ЛПП под действием циркулирующей лецитинхолестеринацилтрансферазы (ЛХАТ) и, наконец, возвращаются в печень. (Модифицировано из Brown MS, Goldstein JL. The hyperlipoproteinemias and other disorders of lipid metabolism. In: Wilson JE, et al., eds. Harrisons principles of internal medicine. 12th ed. New York: McGraw Hill, 1991:1816.) Липопротеинлипаза мышечных клеток и жировой ткани отщепляет от ЛОНП свободные жирные кислоты, которые проникают в клетки, а циркулирующий остаток липопротеина, называемый ремнантным липопротеином промежуточной плотности (ЛПП), содержит в основном эфиры холестерина. Дальнейшие преобразования, которым ЛПП подвергается в крови, ведут к появлению богатых холестерином частиц липопротеинов низкой плотности (ЛНП). Приблизительно 75% циркулирующих ЛНП захватываются печенью и внепеченочными клетками благодаря наличию ЛНП-рецепторов. Остаток подвергается деградации отличными от классического ЛНП-рецепторного пути способами, в основном посредством моноцитарных клеток-мусорщиков. Считается, что холестерин, поступающий в кровь из периферических тканей, транспортируется липопротеинами высокой плотности (ЛВП) в печень, где он вновь включается в липопротеины или секретируется в желчь (путь, включающий ЛПП и ЛНП, называется обратным транспортом холестерина). Таким образом, ЛВП, видимо, играет защитную роль в отношении отложения липидов в атеросклеротических бляшках. В крупных эпидемиологических исследованиях уровень циркулирующего ЛВП обратно коррелирует с развитием атеросклероза. Поэтому ЛВП часто называют хорошим холестерином в противоположность плохому холестерину ЛНП. Семьдесят процентов холестерина плазмы транспортируется в виде ЛНП, и повышенный уровень ЛНП тесно коррелирует с развитием атеросклероза. В конце 1970-х гг. докторами Брауном и Гольдштейном была продемонстрирована центральная роль рецептора ЛНП в доставке холестерина к тканям и его клиренсе из кровотока. Экспрессия рецепторов ЛНП регулируется механизмом отрицательной обратной связи: нормальный или высокий уровень внутриклеточного холестерина подавляет экспрессию рецептора ЛНП на уровне транскрипции, в то время как снижение внутриклеточного холестерина повышает экспрессию рецептора с последующим увеличением захвата ЛНП клеткой. Пациенты с генетическими дефектами рецептора ЛНП (обычно гетерозиготы с одним нормальным и одним дефектным геном, кодирующим рецептор) не могут эффективно удалять ЛНП из кровотока, что приводит к высокому уровню ЛНП в плазме и склонности к преждевременному развитию атеросклероза. Это состояние называется семейной гиперхолестеринемией. Гомозиготы с полным отсутствием рецепторов ЛНП встречаются редко, но у этих людей инфаркт миокарда может развиваться уже в первое десятилетие жизни. Недавно на основании различий в плотности и плавучести были идентифицированы подклассы ЛНП. Лица с более мелкими и плотными частицами ЛНП (свойство, определяемое как генетическими, так и внешними факторами) подвержены более высокому риску инфаркта миокарда, чем обладатели менее плотных разновидностей. Пока остается неясным, почему более плотные частицы ЛНП сопряжены с большим риском, однако это может быть связано с большей подверженностью плотных частиц окислению, ключевому моменту атерогенеза, о чем будет сказано ниже. Возрастает число свидетельств того, что триглицериды сыворотки, в основном транспортирующиеся в составе ЛОНП и ЛПП, могут также играть важную роль в развитии атеросклеротических поражений. Пока не ясно, является ли это их прямым действием или объясняется тем, что уровень триглицеридов обычно находится в обратном соотношении с уровнем ЛВП. Сахарный диабет, начинающийся во взрослом возрасте, является одним из частых клинических состояний, ассоциированных с гипертриглицеридемией и низким уровнем ЛВП, а часто — и с ожирением и артериальной гипертензией. Этот набор факторов риска, который может быть связан с инсулинорезистентностью (обсуждается в главе 13), особенно атерогенен. Читать далее: Преимущества лечения дислипидемии |


Источник