В эритроцитах млекопитающих синтез гемоглобина
Содержание статьи
Гистология.RU
Материал взят с сайта www.hystology.ru
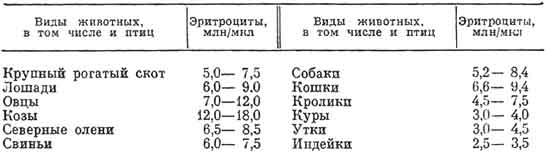
Эритроциты (erythros — красный) — высокоспециализированные клетки, приспособленные для выполнения основной функции крови — транспорта кислорода и углекислого газа в организме. В 1 мкл крови у позвоночных содержится несколько миллионов эритроцитов, а у большинства сельскохозяйственных животных от 5 до 10 млн (табл. 1).
1. Количество эритроцитов в крови животных
Определение количества эритроцитов в крови — важная составная часть общего клинического анализа крови животных, оно проводится либо с помощью счетной камеры, либо в электронных автоматических счетчиках. Количество эритроцитов в крови зависит от вида, породы, возраста животных и может изменяться под влиянием различных факторов — физической нагрузки, барометрического давления, а также при болезнях.
Утратив в процессе развития ядро, зрелые эритроциты у млекопитающих являются безъядерными клетками и имеют форму двояковогнутого круглого диска со средним диаметром круга5 — 7 мкм. Эритроциты крови верблюда и ламы овальной формы. Дисковидная форма увеличивает общую поверхность эритроцита в 1,64 раза по сравнению с поверхностью шара такого же диаметра, что способствует ускорению проникновения кислорода в эритроцит. Эритроциты других позвоночных — птиц, рептилий, амфибии и рыб — овальной формы, имеют ядро с сильно конденсированным хроматином. Они крупнее эритроцитов млекопитающих (например, у саламандр величина их превышает в 100 раз).
В большинстве случаев между количеством эритроцитов и их величиной можно обнаружить обратную зависимость: например, у коз в 1 мкл крови 14 млн эритроцитов, диаметр эритроцита 4 мкм; у лягушки в 1 мкл крови 0,35 млн эритроцитов, диаметр овального эритроцита по длине 22,8 мкм, а по ширине 15,8 мкм. У животных одного вида все эритроциты почти одинаковой величины и появление в крови эритроцитов другой величины и формы считается признаком патологического процесса.
Эритроциты покрыты оболочкой — плазмолеммой (толщиной около 6 нм), содержащей 44% липидов, 47% белков и 7% углеводов. Многие мембранные белки эритроцитов являются гликопротеидами и гликолипидами, их поверхностные концевые олигосахаридные компоненты определяют групповые свойства крови. Мембрана эритроцитов легко проницаема для газов, анионов, обеспечивает активный перенос ионов натрия, облегченный транспорт глюкозы. Внутреннее коллоидное содержимое эритроцитов на 34% состоит из гемоглобина — уникального сложного окрашенного соединения — хромопротеида, в небелковой части которого (теме) имеется двухвалентное железо, способное образовывать особые непрочные связи с молекулой кислорода. Именно благодаря гемоглобину осуществляется дыхательная функция эритроцитов. При высокой концентрации кислорода, особенно в капиллярах легких, происходит присоединение молекул кислорода к атомам железа — образуется оксигемоглобин.
При низкой концентрации кислорода в капиллярах других органов связи между кислородом и железом легко разрываются и кислород отсоединяется — образуется восстановленный гемоглобин, придающий венозной крови синевато-вишневый цвет. Таким образом, функционирование эритроцитов осуществляется непосредственно в сосудистой крови. Обладая большой суммарной поверхностью, эритроциты, кроме транспортировки газов, участвуют в переносе адсорбированных на их оболочке различных веществ — аминокислот, ферментов и др.
Наличие гемоглобина в эритроцитах обусловливает выраженную оксифилию их при окраске мазка крови по Романовскому — Гимзе (смесь кислого красителя эозина и основного — азура II). Эритроциты при этом окрашиваются в красный цвет эозином. Так как эритроциты имеют форму двояковогнутого диска, то центральная часть клетки окрашивается слабее, чем периферическая. Нормальными по окраске считаются эритроциты, центральная часть которых составляет около трети диаметра эритроцита. При некоторых формах анемий центральная бледно окрашенная часть эритроцитов увеличена — гинохромные эритроциты. При суправитальном окрашивании капли крови бриллиантовым крезиловым синим в приготовленном затем мазке можно обнаружить молодые формы эритроцитов, содержащие зернисто-сетчатые структуры. Такие клетки называют ретикулоцитами, они являются непосредственными предшественниками зрелых эритроцитов. При электронной микроскопии в содержимом ретикулоцитов обнаружены компоненты белоксинтезирующей системы — рибосомы, элементы эндоплазматической сети, митохондрии. Радиоавтографическим методом доказано, что в ретикулоцитах продолжает осуществляться синтез белка гемоглобина. Подсчет ретикулоцитов используется для получения информации о скорости образования эритроцитов. Обнаружение в большем количестве по сравнению с нормой ретикулоцитов в крови — признак усиленного эритропоэза.
В зрелых эритроцитах при электронной микроскопии не удается выявить никаких органелл, их внутреннее содержимое имеет высокую электронную плотность.
В период между 100 — 130 днями (у кроликов через 45 — 60 дней) после выхода из красного костного мозга эритроциты фагоцитируются макрофагами главным образом в селезенке, печени и красном костном мозге. При этом от гема гемоглобина отщепляется железо, а из оставшейся части молекулы образуются пигменты желчи, мочи и кала. Железо с помощью белка плазмы крови транспортируется главным образом в костный мозг, где вновь используется при синтезе гемоглобина развивающимися эритроцитами. Таким образом, железо, освобождающееся из фагоцитированных эритроцитов, почти полностью доступно для реутилизации. Избыток железа аккумулируется в макрофагах селезенки или печени в виде гранул гемосидерина, которые выявляют гистохимическими методами.
Эритроциты обладают свойством противостоять различным разрушительным воздействиям — осмотическим, механическим и др. При значительных изменениях концентрации солей в окружающей среде, например при помещении крови в гипотонический раствор, эритроциты набухают, приобретают сферическую форму; мембрана перестает удерживать гемоглобин, и он выходит в окружающую жидкость — явление гемолиза. Выход гемоглобина из эритроцитов может происходить в организме при действии змеиного яда, токсинов, выделяемых некоторыми бактериями, возбудителями паразитарных болезней. Гемолиз развивается также при переливании несовместимой по группе крови. Практически важно при введении в кровь животным жидкостей осуществлять контроль за тем, чтобы вводимый раствор был изотоническим.
У эритроцитов по сравнению с плазмой и лейкоцитами крови относительно большая плотность (удельный вес). Если кровь, предварительно обработанную противосвертывающими веществами, поместить в какой-либо сосуд, то отмечают оседание эритроцитов. Скорость оседания эритроцитов (СОЭ) уживотных разного возраста, пола и вида неодинакова. Высокая СОЭ у лошадей и, наоборот, низкая у крупного рогатого скота. Изменения СОЭ, наблюдаемые в патологии, имеют диагностическое и прогностическое значение.
Отзывов (0)
Добавить отзыв
Источник
Гемоглобин
Гемоглобин (от др.-греч. Гемо — кровь и лат. globus — шар) — это сложная белковая молекула внутри красных клеток крови — эритроцитов (у человека и позвоночных животных). Гемоглобин составляет примерно 98% массы всех белков эритроцита.
Гемоглобин (от др.-греч. Гемо — кровь и лат. globus — шар) — это сложная белковая молекула внутри красных клеток крови — эритроцитов (у человека и позвоночных животных). Гемоглобин составляет примерно 98% массы всех белков эритроцита. За счет своей структуры гемоглобин участвует в переносе кислорода от легких к тканям, и оксида углерода обратно.
Строение гемоглобина
Гемоглобин состоит из двух цепей глобина типа альфа и двух цепей другого типа (бета, гамма или сигма), соединенными с четырьмя молекулами гемма, содержащего железо. Структура гемоглобина записывается буквами греческого алфавита: α2γ2.
Обмен гемоглобина
Гемоглобин образуется эритроцитами в красном костном мозге и циркулирует с клетками в течение всей их жизни — 120 дней. Когда селезенкой удаляются старые клетки, компоненты гемоглобина удаляются из организма или поступают обратно в кровоток, чтобы включиться в новые клетки.
Типы гемоглобина
К нормальным типам гемоглобина относится гемоглобин А или HbA (от adult — взрослый), имеющий структуру α2β2, HbA2 (минорный гемоглобин взрослого, имеющий структуру α2σ2 и фетальный гемоглобин (HbF, α2γ2. Гемоглобин F — гемоглобин плода. Замена на гемоглобин взрослого полностью происходит к 4-6 месяцам (уровень фетального гемоглобина в этом возрасте менее 1%). Эмбриональный гемоглобин образовывается через 2 недели после оплодотворения, в дальнейшем, после образования печени у плода, замещается фетальным гемоглобином.
| Тип гемоглобина | Процент содержания у взрослого человека |
| HbA — взрослый гемоглобин | 98% |
| HbA2 — взрослый гемоглобин минорный | Около 2% |
| HbFi — фетальный гемоглобин | 0,5-1% |
| Эмбриональный гемоглобин | нет |
| HbA1C — гликированный гемоглобин |
Аномальных гемоглобинов более 300, их называют по месту открытия.
Функция гемоглобина
Основная функция гемоглобина — доставка кислорода от легких к тканям и углекислого газа обратно.
Формы гемоглобина
- Оксигемоглобин — соединение гемоглобина с кислородом. Оксигемоглобин преобладает в артериальной крови, идущей от легких к тканям. Из-за содержания оксигемоглобина артериальная кровь имеет алый цвет.
- Восстановленный гемоглобин или дезоксигемоглобин (HbH) — гемоглобин, отдавший кислород тканям
- Карбоксигемоглобин — соединение гемоглобина с углекислым газом. Находится в венозной крови и придает ей темный вишневый цвет.
Как же это происходит? Почему в легких гемоглобин забирает, а в тканях отдает кислород?
Эффект Бора
Эффект был описан датским физиологом Христианом Бором https://en.wikipedia.org/wiki/Christian_Bohr (отцом знаменитого физика Нильса Бора).
Христиан Бор заявил, что при большей кислотности (более низкое значение рН, например, в тканях) гемоглобин будет меньше связываться с кислородом, что позволит его отдать.
В легких, в условиях избытка кислорода, он соединяется с гемоглобином эритроцитов. Эритроциты с током крови доставляют кислород ко всем органам и тканям. В тканях организма с участием поступающего кислорода проходят реакции окисления. В результате этих реакций образуются продукты распада, в том числе, углекислый газ. Углекислый газ из тканей переносится в эритроциты, из-за чего уменьшается сродство к кислороду, кислород выделяется в ткани.
Эффект Бора имеет громадное значение для функционирования организма. Ведь если клетки интенсивно работают, выделяют больше СО2, эритроциты могут снабдить их большим количеством кислорода, не допуская кислородного «голодания». Следовательно, эти клетки могут и дальше работать в высоком темпе.
Какой уровень гемоглобина в норме?
В каждом миллилитре крови содержится около 150 мг гемоглобина! Уровень гемоглобина меняется с возрастом и зависит от пола. Так, у новорожденных гемоглобин значительно выше, чем у взрослых, а у мужчин выше, чем у женщин.
Что еще влияет на уровень гемоглобина?
Некоторые другие состояния также влияют на уровень гемоглобина, например, пребывание на высоте, курение, беременность.
Заболевания, связанные с изменением количества или структуры гемоглобина
- Повышение уровня гемоглобина наблюдается при эритроцитозах, обезвоживании.
- Снижение уровня гемоглобина наблюдается при различных анемиях.
- При отравлении угарным газом образуется карбгемоглобин (не путайте с карбоксигемоглобином!), который не может присоединять кислород.
- Под действием некоторых веществ образуется метгемоглобин.
- Изменение структуры гемоглобина называется гемоглобинопатией. Самые известные и частые заболевания этой группы — серповидно-клеточная анемия, бета-талассемия, персистенция фетального гемоглобина. См.гемоглобинопатии на сайте Всемирной организации здравоохранения https://www.who.int/centre/factsheets/fs308/ru/index.html
Знаете ли Вы?
- У беспозвоночных животных гемоглобин растворен в плазме крови.
- В сутки из легких в ткани переносится около 600 литров кислорода!
- Красный цвет крови придает гемоглобин, входящий в состав эритроцитов. У некоторых червей вместо гемоглобина хлорокруорин и кровь зеленая. А у каракатиц, скорпионов и пауков голубая, так как вместо гемоглобина — содержащий медь гемоцианин.
Другие статьи раздела
Распространенный возбудитель инфекций дыхательных путей (фарингиты, синуситы, отиты, бронхиты и пневмонии). Анализы на антитела используются для диагностики инфекции Chlamydophila pneumoniae при длительных инфекциях дыхательных путей.
Mycoplasma pneumoniae — возбудитель пневмонии человека, острых респираторных заболеваний (ОРЗ), заболеваний верхних дыхательных путей (фарингита, бронхита), а также некоторых нереспираторных заболеваний.
Азооспермия (azoospermia) — отсутствие сперматозоидов в эякуляте
Бактерии — одноклеточные микроорганизмы, некоторые из которых могут вызывать заболевания.
Mycoplasma pneumoniae (микоплазма пневмонии), Chlamydohpila pneumoniae (хламидофила пневмонии, прежнее название Chlamydia pneumoniae)
Гипофиз — непарная железа внутренней секреции, расположенная на основании головного мозга в костном кармане — гипофизарной ямке турецкого седла. Гипофиз вырабатывает гормоны, оказывающие влияние на работу всего организма — рост и развитие, обмен веществ, половую функцию.
Повышенный рост волос (гирсутизм) может быть следствием не только повышенного уровня андрогенов (см. «гиперандрогения»), но и высокой активности 5-альфа-редуказы в коже (фермента волосяных фолликулов, превращающего тестостерон в гораздо более активный дигидротестостерон.
По данным ВОЗ (Всемирная организация здравоохранения) заболеваемость в России составляет более 50 человек на 100 000 населения. Имеет важное значение то, что у женщин значительно чаще чем у мужчин (50-90% против 10%) возможно бессимптомное течение заболевания.
Делеция (ген.) — вид хромосомных мутаций, при котором происходит потеря какого-либо участка хромосомы.
Механизм обратной связи — система, которая используется организмом для контроля некоторых функций и поддержания состояния постоянства организма. Механизм обратной связи использует один из продуктов пути обмена веществ, обычно конечный продукт, для контроля активности пути обмена веществ и регуляции количества этого продукта. Обратная связь может быть отрицательной и положительной.
Источник
Медицинская биохимия, принципы измерительных технологий в биохимии, патохимия, диагностика, биохимия злокачественного роста. Часть 2.
Особенности структурной организации мембраны эритроцитов
Эритроцит окружен плазматической мембраной, структура которой хорошо изучена, идентична таковой в других клетках. Цитоплазматическая мембрана эритроцитов включает бислой фосфолипидов, в то время как белки или «плавают» на поверхности мембран, или пронизывают липиды, обеспечивая прочность и вязкость мембран. Площадь мембраны одного эритроцита составляет около 140 мкм2.
На долю белков приходится примерно 49 %, липидов — 44 %, углеводов -7 %. Углеводы химически связаны либо с белками, либо с липидами и образуют соответственно гликопротеиды и гликолипиды.
Важнейшими компонентами мембраны эритроцитов являются липиды, включающие до 48 % холестерина, 17-28 % — фосфотидилхолина, 13-25 % — сфингомиелина и ряд других фосфолипидов.
Фосфотидилхолин мембраны эритроцитов несет нейтральный заряд, практически не вступает в реакции взаимодействия с положительно заряженными каналами Са2+,, обеспечивая тем самым атромбогенность эритроцитов. Благодаря таким свойствам, как текучесть, пластичность, эритроциты способны проходить через капилляры диаметром ~ 3 мкм.
Белки мембраны эритроцита делят на периферические и интегральные. К периферическим белкам относят спектрин, анкирин, белок 4.1., белок р55, адуцин и др. В группу интегральных белков входит фракция 3, а также гликофорины А, В, С, О, Е. Анкирин образует соединение с р-спектрином. В составе эритроцитов обнаружено около 340 мембранных и 250 растворимых белков.
Пластичность эритроцитов связана с фосфорилированием мембранных белков, особенно белков полосы 4.1.
Белок фракции 4.2. — паллидин обеспечивает связывание спектрин-актин-анкиринового комплекса с фракцией 3, относится к группе трансглутаминазных протеинов.
К числу сократительных белков мембраны эритроцитов относятся р-актин, тропомодулин, строматин и тропомиозин.
Гликофорины — интегральные белки мембраны эритроцитов, определяющие отрицательный заряд, способствующий отталкиванию эритроцитов друг от друг и от эндотелия сосуда.
Протеин 3 — основной белок актинов, регулирующий дефосфорилируемость эритроцита.
Как указывалось выше, мембрана эритроцита представляет собой сложный комплекс, включающий определенным образом организованные липиды, белки и углеводы, которые формируют наружный, средний и внутренний слои эритроцитарной мембраны.
Касаясь пространственного расположения различных химических компонентов эритроцитарной мембраны, следует отметить, что наружный слой образован гликопротеидами с разветвленными комплексами олигосахаридов, которые являются концевыми отделами групповых антигенов крови. Липидным компонентом наружного слоя являются фосфатидилхолин, сфингомиелин и неэстерифицированный холестерин. Липиды наружного слоя мембраны эритроцита играют важную роль в обеспечении постоянства структуры мембраны, избирательности ее проницаемости для различных субстратов и ионов. Вместе с фосфолипидами холестерин регулирует активность мембранно-связанных ферментов путем изменения вязкости мембраны, а также участвует в модификации вторичной структуры ферментов. Молярное отношение холестерин / фосфолипиды в мембранах клеток у человека и многих млекопитающих равно 0,9. Изменение этого соотношения в сторону увеличения наблюдается в пожилом возрасте, а также при некоторых заболеваниях, связанных с нарушением холестеринового обмена.
Снижение текучести мембраны эритроцита и изменение ее свойств отмечается также и при увеличении содержания сфингомиелина,
Средний бислой мембраны эритроцита представлен гидрофобными «хвостами» полярных липидов. Липидный бислой обладает выраженной текучестью, которая обеспечивается определенным соотношением между насыщенными и ненасыщенными жирными кислотами гидрофобной части бислоя. Интегральные белки, к которым относятся ферменты, рецепторы, транспортные белки, обладают активностью только в том случае, если находятся в гидрофобной части бислоя, где они приобретают необходимую для активности пространственную конфигурацию. Поэтому любые изменения в составе липидов эритроцитарной мембраны сопровождаются изменением ее текучести и нарушением работы интегральных белков.
Внутренний слой мембраны эритроцита, обращенный к цитоплазме, состоит из белков спектрина и актина. Спектрин является специфическим белком эритроцитов, его гибкие вытянутые молекулы, связываясь с микрофиламентами актина и липидами внутренней поверхности мембраны, формируют своеобразный скелет эритроцита. Небольшой процент липидов во внутреннем слое мембраны эритроцита представлен фосфатидилэтаноламином и фосфатидилсерином. От наличия спектрина зависит подвижность белков, удерживающих двойной бисой липидов.
Одним из важных гликопротеинов является гликофорин, содержащийся как на внешней, так и на внутренней поверхностях мембран эритроцитов. Гликофорин в своем составе содержит большое количество сиаловой кислоты и обладает значительным отрицательным зарядом. В мембране он располагается неравномерно, образует выступающие из мембраны участки, которые являются носителями иммунологических детерминант.
Строение и состояние эритроцитарной мембраны, низкая вязкость нормального гемоглобина обеспечивают значительные пластические свойства эритроцитам, благодаря которым эритроцит легко проходит по капиллярам, имеющим вдвое меньший диаметр, чем сама клетка, и может принимать самые разнообразные формы. Другим периферическим мембранным белком эритроцитов является анкирин, образующий соединение с молекулой Р-спектрина.
Функции эритроцитарной мембраны
Мембрана эритроцитов обеспечивает регуляцию электролитного баланса клетки за счет активного энергозависимого транспорта электролитов или пассивной диффузии соединений по осмотическому градиенту.
В мембране эритроцитов имеются ионно-проницаемые каналы для катионов Na+, K+, для O2, CO2, Cl- HCO3-.
Транспорт электролитов через эритроцитарную мембрану и поддержание его мембранного потенциала обеспечивается энергозависимыми Na+, K+, Ca2+ — АТФ-азными системами.
Мембрана эритроцитов хорошо проницаема для воды при участии так называемых белковых и липидных путей, а также анионов, газообразных соединений и плохо проницаема для одновалентных катионов калия и натрия.
Белковый путь трансмембранного переноса воды обеспечивается при участии пронизывающего мембрану эритроцитов белка «полосы 3», а также гликофорина.
Молекулярная природа липидного пути переноса воды через эритроцитарную мембрану практически не изучена. Прохождение молекул небольших гидрофильных неэлектролитов через эритроцитарную мембрану осуществляется также, как и перенос воды, за счет белкового и липидного путей. Перенос мочевины и глицерина через мембрану эритроцита обеспечивается за счет ферментативных реакций.
Характерной особенностью мембраны эритроцитов является наличие мощной системы активного транспорта для одновалентных анионов (хлора и фтора), и двухвалентных анионов (SO42-, PO42-) за счет белков — переносчиков.
Перенос органических анионов через эритроцитарную мембрану обеспечивается, как и транспорт неорганических анионов, при участии белка «полосы 3».
Эритроцитарная мембрана обеспечивает активный транспорт глюкозы, кинетика которого обеспечивается зависимостью Михаэлиса-Ментен. Важная роль в транспорте глюкозы через эритроцитарную мембрану отводится полипептиду полосы 4,5 (белки с ММ 55 кД — возможные продукты распада полипептида полосы 3). Высказывается предположение о наличии специфического липидного окружения у белков — переносчиков сахаров в эритроцитарной мембране.
Неравномерное распределение моновалентных катионов в системе эритроцит — плазма крови поддерживается при участии энергозависимой Na+-помпы, осуществляющей трансмембранный обмен ионов Na+ эритроцитов на ионы К+ плазмы крови в соотношении 3:2. Кроме указанного трансмембранного обмена Na+/K+, Na+ помпа осуществляет еще, по крайней мере, четыре транспортных процесса: Na+→ Na+ обмен; K+→K+обмен; одновалентный вход ионов Na+, сопряженный с выходом К+.
Молекулярной основой Na+ помпы является фермент Na+, K+ -АТФ-аза — интегральный белок, прочно связанный с мембранными липидами, состоящий из 2х полипептидных субъединиц с ММ 80-100кД.
Транспортная система имеет 3 центра, связывающих ионов Na+, локализованных на цитоплазматической стороне мембраны. С наружной стороны мембраны на транспортной системе имеется 2 центра связывания ионов К+. Важная роль в поддержании высокой активности фермента отводится мембранным фосфолипидам.
Функционирование Са2+-помпы обеспечивается нуклеотидами, а также макроэргическими соединениями, преимущественно АТФ, ЦТФ, ГТФ, в меньшей степени ГТФ и ЦТФ.
Как в случае Nа+-помпы, функционирование Са2+помпы в эритроцитах связано с проявлениями активности Са2+, Mg2+ -АТФ-азы. В мембране одного эритроцита обнаруживается около 700 молекул Са2+, Mg2+ -АТФ-азы.
Наряду с барьерной и транспортной функциями, мембрана эритроцитов выполняет рецепторную функцию.
Экспериментально доказано наличие на мембране эритроцитов рецепторов к инсулину, эндотелину, церулоплазмину, а2-макроглобулину, α- и β-адренорецепторов. На поверхности эритроцитов находятся рецепторы к фибриногену, обладающие достаточно высокой специфичностью. Эритроциты также несут на мембране рецепторы к гистамину, ТхА2, простациклину.
В мембране эритроцитов обнаруживаются рецепторы для катехоламинов, снижающих подвижность жирных кислот липидов мембран эритроцитов, а также осмотическую устойчивость эритроцитов.
Установлена перестройка структуры мембраны эритроцитов под влиянием низких концентраций инсулина, гормона роста человека, простагландинов группы Е и Е2.
В мембранах эритроцитов высока и ц — АМФ активность. При увеличении концентраций в эритроцитах ц-АМФ ( до 10-6 М) усиливаются процессы фосфорилирования белков, что приводит в свою очередь к изменению степени фосфорилированности и проницаемости мембран эритроцитов для ионов Са2+.
Эритроцитарная мембрана содержит изоантигены различных систем иммунологических реакций, определяющих групповую принадлежность крови человека по этим системам.
Источник